Abstract
Dendritic cells (DC) are professional (specialised) antigen-presenting cells that can capture antigen from apoptotic tumour cells and induce MHC class I- and II-restricted responses. Also, DC fused with tumour cells may be effective for immune response induction. Both cell preparations may be considered as vaccine candidates in a therapeutic approach. We examined autologous T-cell activation by DC that had endocytosed leukaemic B-cell apoptotic bodies (Apo-DC) and compared it to the T-cell stimulatory capacity of DC that were fused with tumour cells. Following incubation, 22.6±6.2 (mean±s.e.m.) of DC had endocytosed leukaemic cells, while the frequency of DC–leukaemic cell hybrids was 10.5±2.6%. Apo-DC and hybrid cells both demonstrated the ability to stimulate a tumour-specific T-cell immune response in vitro. A T-cell proliferation response was also observed in four out of five CLL patients when using Apo-DC. However, fusion hybrids lacked the ability to elicit a proliferative response. Apo-DC also induced an IFN-γ response, as did hybrid cells. The cytokine response induced by Apo-DC was significantly higher than that induced by fusion (P<0.05). This study shows that endocytosed apoptotic tumour cells induced a significantly stronger T-cell response than DC hybrids; and as such should be a better candidate for vaccine production.
Similar content being viewed by others

Introduction
The slow, smouldering disease presented initially in many of B-chronic lymphocytic leukaemia (B-CLL) patients makes this disease an ideal candidate for immune therapeutic intervention. Active immunotherapy against leukaemia-associated antigens requires the transfer of tumour antigens from malignant cells to ‘professional’ antigen-presenting cells (APC) to initiate a clinically relevant antitumour response. Presentation of antigens by APC is necessary as tumour cells themselves usually lack the accessory molecules required for the induction and expansion of cytolytic T-cells that recognise tumour antigens in MHC class I- and II-restricted manner.1
Dendritic cells (DC) possess the unique ability to efficiently present antigens to naïve T-cells and are a key player in the primary immune response.2,3 DC loaded with immunoglobulin VH-CDR3 (complementarity determining region III) peptides could elicit MHC class I- and II dependent immune response in B-CLL.4 In animal models, fusion of DC with tumour cells induced a polyclonal antitumour immune response leading to the elimination of metastatic tumour.5
Another approach to induce immune response is to use DC pulsed with apoptotic bodies derived from tumour cells. MHC class I and II molecules of DC present proteins better from phagocytosed cellular fragments compared to preprocessed peptides.6 DC that had acquired antigens from apoptotic bodies induced MHC class I-restricted CTL and antitumour immunity in vivo.7,8
In this study, we compare the efficacy of DC that have endocytosed apoptotic bodies or have been fused with tumour cells as APC to stimulate autologous T-cell responses against CLL.
Materials and methods
Patients
All research samples were collected in accordance with the Helsinki declaration on human research and as per the protocol approved by the institutional ethics committee. Six B-CLL patients (three males and three females) with a mean age of 68.5 (range 50–80 years) were studied (Table 1). Three healthy control donors (one male and two females with a mean age of 53 years (range 42–60) were also included. The diagnosis and staging (Rai) as well as the criteria for progressive and nonprogressive disease have been described earlier.9,10 Four patients had stage 0, one patient stage 3 and one patient stage 4 disease. All patients had nonprogressive disease and had not previously received any therapy.
Cell separation
Peripheral blood was collected by venipuncture in sterile heparinised tubes. Peripheral blood mononuclear cells (PBMC) were obtained by centrifugation on a Ficoll-paque gradient (Amersham Pharmacia Biotech, Uppsala, Sweden) as described.11
B-cell purification
PBMC were washed three times with phosphate-buffered saline (PBS), placed on nylon wool columns (Biotest, Breiech, Germany) and B cells were eluted from the column.12 Effluent B cells were collected and cell purity was determined by flow cytometry.
T-cell purification
PBMC of patients were washed three times with PBS and passed through a nylon wool column (Biotest). The effluent cells were further enriched immunomagnetically with anti-CD19 MidiMACS beads (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's specification. The purity of T-cells was >95% as determined by flow cytometry after anti-CD3 staining.
Flow cytometry and immunophenotyping
Cells were analysed by flow cytometry (FACScan Becton-Dickinson (BD), Mountain View, CA, USA) using conjugated CD3, CD5, CD14, CD19 and CD83 monoclonal antibodies (MAb), and relevant isotype controls CD19 and CD83 antibodies were obtained from DAKO A/S (Glostrup, Denmark).
Surface marker staining was performed as described earlier.13 Intracellular staining was carried out according to a modified method as described by Mendes et al.14 Briefly, DC were fixed in 0.5 ml of fixing solution (2% paraformaldehyde in PBS). After washing, the cells were permeabilised with 0.5 ml of 1 × permeabilising solution (BD) and incubated for 15 min followed by washing in buffer (0.1% saponin, 0.2% bovine serum albumin and 0.1% sodium azide). Anti-CD19 (10 μl) or a relevant isotype control antibody was added. The cells were incubated for an additional 30 min in dark and then washed twice with washing buffer and once with PBS. The mean fluorescence intensity and the relative cell frequency expressing the respective surface markers were analysed using the CELLQuest software (BD).
Generation of DCs
DC were generated from CD14+ cells as previously described.15 Briefly, PBMC incubated with anti-CD14+-coated MiniMACS beads (Miltenyi) were passed through MidiMACS columns according to the manufacturer's instruction. CD14+ cells were resuspended in the medium and purity was determined by flow cytometry (83.2±3.8%; mean±s.e.m.). The isolated CD14+ cells were cultured with recombinant (r) GM-CSF (50 ng/ml) (Schering-Plough Research Institute (SPRI), Kenilworth, NJ, USA) and rIL-4 (SPRI; 10 ng/ml) for 5 days. For maturation of DC, rTNF-α (Pharmingen, San Diego, CA, USA) (20 ng/ml) was added at day 5 and the cells were cultured for further 2 days. The harvested cells had the typical morphology of DC with CD3−, CD14−, CD19−, CD83+, CD80+.13
Fusion of tumour B cells and DC (hybrids)
Fusion was performed according to a modified method by Gong et al.16 To optimise the time and ratio of tumour cells:DC, different ratios and times were tested. The ratio of 10:1 tumour and DC fused for 5 min was determined to be optimum. Autologous DC and B cells were washed twice with serum-free medium and incubated for 5 min at a ratio of 1:10 in 50% polyethylene glycol (PEG) solution (Sigma-Aldrich, UK). Prewarmed medium (37°C) was added slowly to the fused cells to dilute the PEG, followed by centrifugation. After washing, the cells were resuspended in the medium and analysed by flow cytometry. DC with CD83+ and CD19+ surface markers were considered as hybrid cells.
Generation of Apo-DC
After purification of B cells, induction of apoptosis was performed according to a modified method by Comby et al.17 Briefly, cells were subjected to 5 Gy irradiation from a Cobalt source (Alcyon, CGR, Paris, France). Irradiated cells were resuspended in the medium and incubated at 37°C with 5% CO2 for 24 h. The rate of apoptosis was analysed using annexin-V FITC apoptosis detection kit (Annexins Research, PharMingen, BD, USA) and 7AAD (PharMingen, BD) using flow cytometry.1 At the end of 24 h of culture, apoptotic cells were spun down, washed and added at a ratio of 1:2 to immature DC. The cells were cocultured for an additional 24 h. Following the initial culture to allow endocytosis of apoptotic bodies, TNFα (20 ng/ml) was added and cultured for an additional 36 h for maturation of DC. After maturation of DC the engulfment of apoptotic cells was measured by intracellular staining of DC for B-cell marker (CD19) along with surface staining for DC marker (CD83). Cells with CD83 and CD19 markers as gated for DC were considered as DC with apoptotic bodies designated Apo-DC.
T-cell proliferation assay
The ability of hybrid cells and Apo-DC to induce an autologous T-cell response was determined in an in vitro proliferation assay. Fusion hybrids or Apo-DC (2 × 104) were added to 2 × 105 autologous T-cells in a 96-well U-bottom plate and incubated at 37°C for 5 days in RPMI 1640 medium (Gibco, BRL, UK) with 10% heat-inactivated pooled human AB+ serum, 1 mM L-glutamine, 100 IU/ml penicillin and 100 μg/ml streptomycin (referred to as medium). Cultures were incubated for a total of 5 days, with 1 μCi [3H]thymidine (Amersham Pharmacia Biotech, Uppsala, Sweden) added to each well for the final 18 h. Cells were harvested and the incorporated radioactivity was measured in a beta-scintillation counter (Microbeta 1450, Wallac, Turku, Finland). Proliferation of T-cells alone or cocultured with autologous DC in the absence of tumour antigen was examined as controls. Baseline proliferation values were determined by assaying T-cell stimulation by Apo-DC and B-cell–DC fusion hybrids of three healthy donors.
Enzyme-linked immunospot assay (ELISPOT)
Identification of IFN-γ secreting T-cells was performed as described earlier.18 The 96-well flat-bottomed nitrocellulose microtiter plates (Millipore AB, Stockholm, Sweden) were coated with anti-human IFN-γ MAb (clone 1-D1 K MABTECH Sweden) at 4°C overnight. Fused cells or Apo-DC (2 × 104) were added to 2 × 105 autologous T-cells and incubated at 37°C overnight. They were then transferred to the washed microtiter plates and incubated at 37°C for 1 day. Biotinylated anti-IFN-γ avidin-conjugated alkaline phosphatase and BCIP® (Sigma Diagnostic, St Louis, MO, USA) were used for the colorimetric reaction. Spots were quantitated using an automatic ELISPOT reader (Axioplan2 Microscope reader, Carl Zeiss, Jena, Germany).
Values obtained with autologous T-cells from three nonleukaemic healthy donors cocultured with fusion or Apo-DC were used to define the baseline value for IFN-γ secreting T-cells.
Results
Immunophenotyping
The process of immunomagnetic selection allowed us to obtain cell populations of high purity at the start of the culture. The purity of CD19+ B-cells in the six B-CLL patients ranged from 90 to 98% (95.3±1.3; mean±s.e.m.). The purity of monocytes (CD14+) ranged from 70 to 99% (83% ±3.8; mean±s.e.m.).
DC generated by the method described in the Materials and methods section were nonadherent and displayed the characteristic morphology described for DC. They also displayed the typical phenotype with high CD83 expression and lack of CD14 (<1% were CD14+) in CLL patients as well as in healthy control donors.
Quantitation of DC–tumour hybrids was performed by calculating the percentage of cells showing dual positivity for DC marker (CD83) and B-cell marker (CD19). By this method, an average hybridisation frequency of 10.5±2.6% (mean±s.e.m.) was estimated over five experiments (Table 2). We speculate that the actual percentage of hybrids generated may have been slightly higher, since dilution of the relevant surface markers over two cells following fusion of their membranes may have resulted in lowered detection sensitivity.
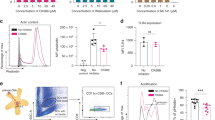
Following irradiation, 49.2±9.8% (mean±s.e.m.) of the B-CLL cells were determined to be apoptotic as measured by Annexin-V positivity. Following coincubation with DC, 22.6 ±6.2% (mean±s.e.m.) of CD83+ cells was determined to have endocytosed CD19+ apoptotic bodies (Figure 1). Thus, in a comparison between fusion hybrids and apoptotic B-CLL cells, Apo-DC had a greater uptake of CLL antigens and thus represented a more efficient antigen delivery system than DC–tumour fusions. In preliminary experiments, irradiated or unirradiated CLL cells by themselves did not demonstrate any T-cell stimulatory activity (data not shown).

Phenotypic markers expressed by DC following endocytosis of apoptotic B-CLL cells. The histogram shows fluorescence value of cells, gated for the forward and side scatter characteristic of DC. The fraction of cells, dual positive for CD83-FTIC and CD19-PE, represents DC that had endocytosed apoptotic B-CLL cells (Apo-DC). Data shown were obtained with one patient and similar results were noted with cells of the other patients.
Proliferative T-cell response to tumour hybrids and Apo-DC
The capacity to stimulate proliferation of unprimed T-cells is an essential criterion for an APC to generate antileukaemic immune response. The ability of fused and Apo-DC to induce an autologous T-cell response was measured in an in vitro proliferation assay. Average proliferation value, obtained with Apo-DC from three healthy nonleukaemic volunteers, has been depicted as the broken line. Proliferation of autologous T-cells by fusion hybrids from normal nonleukaemic donors was lower than the value represented by the broken line (data not shown). No significant proliferative response was observed in patients or in control donors when DC fusion hybrids were used as stimulators. However, autologous T-cells from all five patients showed a proliferative response with Apo-DC (Figure 2).

Proliferation of autologous T-cells following stimulation with DC fusion hybrids or Apo-DC. Fusion hybrids and Apo-DC of B-CLL patients listed in Table 1 were cocultured with purified autologous T-cells as described in Materials and methods. [3H]thymidine was added for the final 18 h of culture. Results represent mean±s.d. c.p.m. of triplicate cultures. Broken horizontal line represents the baseline proliferation obtained with Apo-DC and autologous T-cells of three nonleukaemic donors (enriched normal B-cells). Comparable or lower value than that depicted by the baseline was obtained when fusion hybrids and autologous T-cells of three nonleukaemic donors were tested (data not shown).
Production of IFN-γ by autologous T-cells in response to stimulation by DC–tumour hybrids and Apo-DC
Autologous T-cells stimulated with DC–tumour fusion hybrids or Apo-DC were analysed for IFN-γ production by the ELISPOT assay. DC–tumour fusion hybrids and Apo-DC elicited IFN-γ responses in three of five and four of four patients respectively (Figure 3). IFN-γ secretion was statistically significantly higher (P<0.05) using Apo-DC as compared to hybrid tumour cells. T-cells from nonleukaemic donors stimulated with autologous Apo-DC yielded <10 spots/106cells (n=3), which was considered as the baseline value (broken line). Comparable or lower values were also obtained when autologous fusion hybrids of nonleukaemic donors were used as APC (data not shown).

Gamma interferon release of autologous T-cells after stimulation with fusion hybrids or Apo-DC. T-cells cocultured with fusion hybrids or Apo-DC were assayed for IFN-γ release by the ELISPOT assay. Data represent mean±s.d. from triplicate wells. Baseline value represented by the broken line was determined using T-cells cocultured with autologous Apo-DC from three nonleukaemic donors (enriched normal B cells). Comparable or lower value than that depicted by the baseline was obtained when fusion hybrids and autologous T-cells of three nonleukaemic donors were tested (data not shown). Purified T-cells from the same patients, polyclonally stimulated with phytohaemagglutinin (PHA) was included as a positive control (data not shown). ND indicates that Apo-DC were not tested for patient 1.
Discussion
The use of DC as cellular adjuvants to elicit antitumour immune responses is an important development in the area of cancer vaccination. In this study we observed that DC primed with apoptotic tumour cells and, to a lesser extent, fusion hybrids of DC and B-CLL cells stimulate autologous T-cell responses. Our observations confirm the existence of immunodominant epitopes on tumour cells which are capable of eliciting autologous immune responses when presented in conjunction with potent APC. A comparison of DC–tumour hybrids and Apo-DC revealed that both IFN-γ production and autologous T-cell proliferative responses are induced when apoptotic bodies were used, whereas only IFN-γ production was noted with DC–tumour cell hybrids.
The existence of naturally occurring leukaemic cell and tumour IgVH-CDR3-specific T-cells in B-CLL4,15,19,20 as well as in multiple myeloma and B-cell lymphoma21,22,23 has been reported earlier. Thus, tumour cells exhibit antigenic epitopes capable of inducing a tumour-specific cellular response. Such antigens may potentially be used in a therapeutic approach to induce a T-cell-specific immunity. Multiple myeloma patients, vaccinated with the complete tumour derived idiotypic immunoglobulin, mounted MHC class I- as well as MHC class II- restricted18 T-cell response directed against VH-CDR1-3 regions.
Apoptotic tumour cells have also been described as a suitable vehicle to deliver antigens for processing and presentation.24 An advantage offered by apoptotic bodies is that presentation of intracellular proteins released into tumour lysates or necrotic cells that are not relevant to induction of antitumour responses can be avoided.25 Apo-DC are an attractive alternative since phagocytosed cellular fragments are 300 times more efficient in forming MHC–peptide complexes than preprocessed peptides.26 Considering the above reports as well as the observations of the present study, apoptotic bodies from leukaemic B-cells engulfed by DC (Apo-DC) may be a good antigen presentation system for the induction of immune response in B-CLL patients.
The internalisation rate of apoptotic cells in our study was 22.6±6.2% (mean±s.e.m.). Hoffmann et al24 reported 27% internalisation after 2 h coincubation and 48% after overnight incubation. An autologous T-cell immune response was observed in five out of six CLL patients using Apo-DC (Figure 3). This is similar to a report showing that phagocytosed apoptotic tumour cells induced T-cell proliferation and IFN-γ production in a mouse melanoma model.27 When the fusion DC–normal B-cells were used as APC, no significant proliferation response was observed in autologous normal donor T-cells. Our observations contrast previous reports that demonstrate autologous T-cell stimulation by DC fusion hybrids in solid tumours.28 The disparate observations may be because of differences in solid tumours and B-CLL cells. In four out of five CLL patients, IFN-γ production was observed but in a lower amount than that induced by Apo-DC (Figure 3). Our findings are similar to those reported with pancreatic tumour cells that demonstrate activation of cytotoxic T-cells following cross priming by apoptotic bodies.29 The reasons for the weak proliferation response in T-cells cocultured with DC fusion hybrids are not clear, but it has been shown that purified hybrid cells are more effective in inducing an immune response (IFN-γ production) as compared to fusion mixture.30 The higher proliferative response that we obtained using Apo-DC may be attributed to the fact that particulate antigens prime T-cell more effectively.31 Furthermore, residual nonhybridised B-CLL cells may serve to anergise T-cells.32 All these parameters may affect the proliferation response when using DC fusion hybrids to stimulate autologous antileukaemic T-cells.
In conclusion, this study provides preclinical data on developing a DC-based vaccination strategy in CLL. Our results demonstrate that it is feasible to produce Apo-DC with relative ease. These cells can be efficiently utilised for generating an antileukaemic immune response in B-CLL patients.
References
Berger CL, Xu AL, Hanlon D, Lee C, Schechner J, Glusac E . Induction of human tumor-loaded dendritic cells. Int J Cancer 2001; 91: 438–447.
Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ . Immunobiology of dendritic cells. Annu Rev Immunol 2000; 18: 767–811.
Biragyn A, Kwak LW . Designer cancer vaccines are still in fashion. Nat Med 2000; 6: 966–968.
Rezvany MR, Jeddi-Tehrani M, Rabbani H, Lewin N, Avila-Carino J, Osterborg A . Autologous T lymphocytes may specifically recognize leukaemic B cells in patients with chronic lymphocytic leukaemia. Br J Haematol 2000a; 111: 608–617.
Gong J, Chen D, Kashiwaba M, Kufe D . Induction of antitumor activity by immunization with fusions of dendritic and carcinoma cells. Nat Med 1997; 3: 558–561.
Rovere P, Vallinoto C, Bondanza A, Crosti MC, Rescigno M, Ricciardi-Castagnoli P . Bystander apoptosis triggers dendritic cell maturation and antigen-presenting function. J Immunol 1998; 161: 4467–4471.
Albert ML, Sauter B, Bhardwaj N . Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature 1998; 392: 86–89.
Henry F, Bretaudeau L, Barbieux I, Meflah K, Gregoire M . Induction of antigen presentation by macrophages after phagocytosis of tumour apoptotic cells. Res Immunol 1998; 149: 673–679.
Rai KR, Sawitsky A, Cronkite EP, Chanana AD, Levy RN, Pasternack BS . Clinical staging of chronic lymphocytic leukemia. Blood 1975; 46: 219–234.
Idestrom K, Kimby E, Bjorkholm M, Mellstedt H, Engstedt L, Gahrton G . Treatment of chronic lymphocytic leukaemia and well-differentiated lymphocytic lymphoma with continuous low- or intermittent high-dose prednimustine versus chlorambucil/prednisolone. Eur J Cancer Clin Oncol 1982; 18: 1117–1123.
Avila-Carino J, Lewin N, Yamamoto K, Tomita Y, Mellstedt H, Brodin B . EBV infection of B-CLL cells in vitro potentiates their allostimulatory capacity if accompanied by acquisition of the activated phenotype. Int J Cancer 1994; 58: 678–685.
Boyum A . Separation of lymphocytes, lymphocyte subgroups and monocytes: a review. Lymphology 1977; 10: 71–76.
Rezvany MR, Jeddi-Tehrani M, Biberfeld P, Soderlund J, Mellstedt H, Osterborg A . Dendritic cells in patients with non-progressive B-chronic lymphocytic leukaemia have a normal functional capability but abnormal cytokine pattern. Br J Haematol 2001; 115: 263–271.
Mendes R, Bromelow KV, Westby M, Galea-Lauri J, Smith IE, O'Brien ME . Flow cytometric visualisation of cytokine production by CD3− CD56+ NK cells and CD3+ CD56+ NK-T cells in whole blood. Cytometry 2000; 39: 72–78.
Rezvany MR, Jeddi-Tehrani M, Rabbani H, Ruden U, Hammarstrom L, Osterborg A . Autologous T lymphocytes recognize the tumour-derived immunoglobulin VH-CDR3 region in patients with B-cell chronic lymphocytic leukaemia. Br J Haematol 2000b; 111: 230–238.
Gong J, Nikrui N, Chen D, Koido S, Wu Z, Tanaka Y . Fusions of human ovarian carcinoma cells with autologous or allogeneic dendritic cells induce antitumor immunity. J Immunol 2000; 165: 1705–1711.
Comby E, Andre I, Troussard X, Lebrun E, Sola B, Ballet JJ . In vitro evaluation of B-CLL cells apoptotic responses to irradiation. Leukaemia Lymphoma 1999; 34: 159–166.
Osterborg A, Yi Q, Henriksson L, Fagerberg J, Bergenbrant S, Jeddi-Tehrani M . Idiotype immunization combined with granulocyte-macrophage colony-stimulating factor in myeloma patients induced type I, major histocompatibility complex-restricted, CD8- and CD4-specific T-cell responses. Blood 1998; 91: 2459–2466.
Buhmann R, Nolte A, Westhaus D, Emmerich B, Hallek M . CD40-activated B-cell chronic lymphocytic leukemia cells for tumor immunotherapy: stimulation of allogeneic versus autologous T-cells generates different types of effector cells. Blood 1999; 93: 1992–2002.
Kato K, Cantwell MJ, Sharma S, Kipps TJ . Gene transfer of CD40-ligand induces autologous immune recognition of chronic lymphocytic leukemia B cells. J Clin Invest 1998; 101: 1133–1141.
Fagerberg J, Yi Q, Gigliotti D, Harmenberg U, Ruden U, Persson B . T-cell-epitope mapping of the idiotypic monoclonal IgG heavy and light chains in multiple myeloma. Int J Cancer 1999; 80: 671–680.
Wen YJ, Lim SH . T-cells recognize the VH complementarity-determining region 3 of the idiotypic protein of B cell non-Hodgkin's lymphoma. Eur J Immunol 1997; 27: 1043–1047.
Wen YJ, Lim SH . In vivo immune responses to idiotypic VH complementarity-determining region 3 peptide vaccination in B-cell non-Hodgkin's lymphoma. Br J Haematol 1998; 103: 663–668.
Hoffmann TK, Meidenbauer N, Dworacki G, Kanaya H, Whiteside TL . Generation of tumor-specific T-lymphocytes by cross-priming with human dendritic cells ingesting apoptotic tumor cells. Cancer Res 2000; 60: 3542–3549.
Henry F, Boisteau O, Bretaudeau L, Lieubeau B, Meflah K, Gregoire M . Antigen-presenting cells that phagocytose apoptotic tumor-derived cells are potent tumor vaccines. Cancer Res 1999; 59: 3329–3332.
Inaba K, Turley S, Yamaide F, Iyoda T, Mahnke K, Inaba M . Efficient presentation of phagocytosed cellular fragments on the major histocompatibility complex class II products of dendritic cells. J Exp Med 1998; 188: 2163–2173.
Chen Z, Moyana T, Saxena A, Warrington R, Jia Z, Xiang J . Efficient antitumor immunity derived from maturation of dendritic cells that had phagocytosed apoptotic/necrotic tumor cells. Int J Cancer 2001; 93: 539–548.
Gong J, Avigan D, Chen D, Wu Z, Koido S, Kashiwaba M . Activation of antitumor cytotoxic T lymphocytes by fusions of human dendritic cells and breast carcinoma cells. Proc Natl Acad Sci USA 2000; 97: 2715–2718.
Schnurr M, Scholz C, Rothenfusser S, Galambos P, Dauer M, Robe J . Apoptotic pancreatic tumor cells are superior to cell lysates in promoting cross-priming of cytotoxic T-cells and activate NK and gammadelta T-cells. Cancer Res 2002; 62: 2347–2352.
Li JLMH, Franek KJ, Burgin KE, Wagner TE, Wei Y . Purified hybrid cells from dendritic cell and tumor cell fusions are superior activators of antitumor immunity. Cancer Immunol Immunother 2001; 50: 445–455.
Speidel K, Osen W, Faath S, Hilgert I, Obst R, Braspenning J . Priming of cytotoxic T lymphocytes by five heat-aggregated antigens in vivo: conditions, efficiency, and relation to antibody responses. Eur J Immunol 1997; 27: 2391–2399.
Kneitz C, Goller M, Wilhelm M, Mehringer C, Wohlleben G, Schimpl A . Inhibition of T-cell/B-cell interaction by B-CLL cells. Leukemia 1999; 13: 98–104.
Acknowledgements
This work was supported by grants from The Swedish Cancer Foundation, The Cancer Society in Stockholm, King Gustav Vth Jubilee Fund, The Karolinska Institutet Foundation, the Swedish Medical Society, and the Swedish Society for Medical Research, Gunnar Nilsson Foundation, and the Cancer and Allergy Foundation, and the Torsten and Ragnar Söderberg Foundation.
The authors gratefully acknowledge Lillemor Lauren for technical assistance.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Kokhaei, P., Rezvany, M., Virving, L. et al. Dendritic cells loaded with apoptotic tumour cells induce a stronger T-cell response than dendritic cell–tumour hybrids in B-CLL. Leukemia 17, 894–899 (2003). https://doi.org/10.1038/sj.leu.2402913
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.leu.2402913
Keywords
This article is cited by
-
Apoptotic blebs from leukemic cells as a preferred source of tumor-associated antigen for dendritic cell-based vaccines
Cancer Immunology, Immunotherapy (2014)
-
Vaccination with dendritic cells loaded with tumor apoptotic bodies (Apo-DC) in patients with chronic lymphocytic leukemia: effects of various adjuvants and definition of immune response criteria
Cancer Immunology, Immunotherapy (2012)
-
Differential effects of Paclitaxel on dendritic cell function
BMC Immunology (2010)
-
Dendritic Cell-Ewing’s Sarcoma Cell Hybrids Enhance Antitumor Immunity
Clinical Orthopaedics and Related Research (2008)
-
Development of a dendritic cell-based vaccine for chronic lymphocytic leukemia
Cancer Immunology, Immunotherapy (2008)
