
Abstract
Aim:
Ginger rhizome is used worldwide as a spicy flavor agent. This study was designed to explore the potential effects of pungent ginger components, 6-, 8-, and 10-gingerol, on human cytochrome P450 (CYP450) enzymes that are responsible for the metabolism of many prescription drugs.
Methods:
The activities of human CYP2C9, CYP2C19, CYP2D6, and CYP3A4 were analyzed using Vivid P450 assay kits. The mRNA expression of CYP3A4 in human hepatocellular carcinoma cell line HepG2 was measured using quantitative real-time PCR assay.
Results:
All three gingerols potently inhibited CYP2C9 activity, exerted moderate inhibition on CYP2C19 and CYP3A4, and weak inhibion on CYP2D6. 8-Gingerol was the most potent in inhibition of P450 enzymes with IC50 values of 6.8, 12.5, 8.7, and 42.7 μmol/L for CYP2C9, CYP2C19, CYP3A4, and CYP2D6, respectively. By comparing the effects of gingerols on CYP3A4 with three different fluorescent substrate probes, it was demonstrated that the inhibition of gingerols on CYP3A4 had no substrate-dependence. In HepG2 cells, 8-gingerol and 10-gingerol inhibited, but 6-gingerol induced mRNA expression of CYP3A4.
Conclusion:
6-, 8-, and 10-gingerol suppress human cytochrome P450 activity, while 8- and 10-gingerol inhibit CYP3A4 expression. The results may have an implication for the use of ginger or ginger products when combined with therapeutic drugs that are metabolized by cytochrome P450 enzymes.
Similar content being viewed by others
Introduction
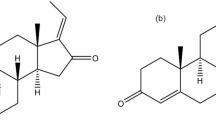
Ginger rhizome (Zingiber officinale Roscoe), commonly known as ginger, is used worldwide as a spicy flavor agent. It is a well-known dietary supplement for treating nausea1 and motion sickness2. Ginger rhizomes are widely consumed in various forms: fresh paste, dried powder, slices preserved in syrup, candy (crystallized ginger), and in tea and beverages. In many countries, particularly those in Southeast Asia, fresh and processed ginger products have a long history of use as traditional medicine for treating a broad range of ailments, such as the common cold, inflammation, rheumatic disorder, and gastrointestinal discomforts3. Ginger contains 1.0% to 3.0% volatile oils and a number of pungent compounds2; 6-, 8-, and 10-gingerol (Figure 1A) are the major pungent constituents of ginger that lead to the spicy taste4, and 6-gingerol is the most abundant one. The gingerols, which are often chemically differentiated by the length of their unbranched alkyl chains, account for the pharmacological activity of ginger5.

Structures of gingerols (A) and Vivid fluorescent probes (B).
The pharmacological properties of gingerols include antioxidant, analgesic and anti-inflammatory6, antipyretic7, cardiotonic8, and hypothermic9 activities. More recent studies have suggested that gingerols exhibit preventive activity against cancers of the skin10, pancreas11, gastrointestinal tract12, colon13, and breast14. Ginger can also effectively prevent nausea induced by cancer treatment15. It has been reported that 6-, 8-, and 10-gingerol exist in ginger-containing dietary supplements, spices, food products and beverages16. Pharmacokinetic studies in humans have shown that after absorption and metabolism, 6-, 8-, and 10-gingerol are present in the serum as glucuronide and sulfate conjugates17.
There is an increasing interest in determining the importance of drug-drug, drug-nutrient, and drug-dietary supplements interactions in basic, translational, and clinic research settings18,19,20. It is necessary to evaluate the potential influence of gingerols on human drug metabolism. The present study was designed to test the hypothesis that gingerols affect human P450 enzymes, and such effects may have an implication in clinical practice when ginger is combined with therapeutic drugs that are metabolized by P450s. In the present study, we investigated the effects of gingerols on several P450 enzymes that have been shown to have pharmacogenomic variations in humans, including CYP2C9, CYP2C19, CYP2D6, and CYP3A4. The potential influence of gingerols on the cellular transcription activity of CYP3A4, the most abundant CYP enzyme in the liver, was also examined.
Materials and methods
Chemicals and reagents
6-, 8- and 10-gingerol were purchased from Phytomarker Ltd (Tianjin, China) with purities of 98.02%, 98.87%, and 99.41%, respectively, as analyzed by HPLC. Miconazole nitrate salt, quinidine, and ketoconazole were purchased from Sigma-Aldrich (St Louis, MO, USA). Sulfaphenazole was a gift from Ms Yuan-yuan DAI at the Cancer Hospital/Institute, Chinese Academy of Medical Sciences (Beijing, China). Chromatographic-grade methanol and acetonitrile were purchased from Tedia Company (Fairfield, OH, USA). Rifampicin (RIF) was purchased from AppliChem GmbH (Darmstadt, Germany).
Determination of P450 activity with Vivid P450 assay kits
The inhibitory effects of gingerols on the catalytic activities of cDNA-expressed human P450 enzymes were determined using Vivid P450 screening kits (Vivid OOMR substrate CYP2C9 red, Vivid EOMCC substrate CYP2C19 blue, Vivid EOMCC substrate CYP2D6 blue, Vivid DBOMF substrate CYP3A4 green, Vivid BOMR substrate CYP3A4 red, and Vivid BOMCC substrate CYP3A4 blue) (Figure 1B) according to the manufacturer's instructions (Invitrogen Corporation; Carlsbad, CA, USA). Each kit contained P450 reaction buffer, P450 BACULOSOMES reagent, a fluorescent substrate, a fluorescent standard, the regeneration system (333 mmol/L glucose-6-phosphate and 30 000 U/L glucose-6-phosphate dehydrogenase in 100 mmol/L potassium phosphate, pH 8.0), and 10 mmol/L NADP+ in 100 mmol/L potassium phosphate, pH 8.0.
In brief, the assays were performed in Costar black-wall 96-well plates with ultra-thin clear bottoms (Corning Inc, Corning, NY, USA) in kinetic assay mode. Stock solutions (10 mmol/L) of gingerols in methanol were prepared and diluted to various concentrations (250, 50, 10, 5, and 1 μmol/L). In each well, 40 μL of the test compound in solution, solvent or a positive inhibitor control was incubated with 50 μL of pre-mixture (containing BACULOSOMES reagent, the regeneration system and reaction buffer) or 50 μL of reaction buffer only as a background control at room temperature (25 °C) for 20 min. The reaction was initiated by the addition of 10 μL/well of a mixture of substrate and NADP+ with a respective concentration of the Vivid substrate (Table 1). The plates were read immediately for fluorescence changes every 30 s at 37 °C for 30 min using a FlexStation II384 fluorometric plate reader (Molecular Devices; Sunnyvale, CA, USA) with appropriate excitation and emission wavelengths for each P450 enzyme (Table 1). The final methanol volume in the reactions was ≤1%. The positive control compounds were sulfaphenazole (for CYP2C9), miconazole nitrate salt (for CYP2C19), quinidine (for CYP2D6), and ketoconazole (for CYP3A4); the final inhibitor concentrations of the inhibitors were chosen according to the Vivid P450 screening kit protocol to produce an inhibition of ≥90%.
Cell culture and treatment
The human hepatocellular carcinoma cell line HepG2 was cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 100 units/mL penicillin and streptomycin at 37 °C with 5% CO2. For the CYP3A4 expression study, 5×105 cells per well were seeded in 6-well plates. After 24 h of incubation, the cells were treated with the tested compounds at a final concentration of 10 μmol/L for 48 h. All compounds were dissolved in sterile DMSO to make 50 mmol/L stock solutions and kept at -20 °C until use; 1% DMSO was used as a vehicle control.
Quantitative real-time PCR analysis
Total RNA was isolated using the TRIzol reagent (Invitrogen) according to the manufacturer's protocol. First-strand cDNA was synthesized using a ReverTra Ace qPCR RT Kit (Toyobo, Japan) with 1 μg of total RNA. Quantitative real-time PCR for CYP3A4 was performed using SYBR Green fluorescent dye methodology as described previously21.
Statistical analysis
The quantitative data were presented as the mean±SD from at least three independent experiments. All the statistical analyses were conducted with Excel software. The statistical significance of differences between the control and treated groups was examined using Student's t test. A P value <0.05 was considered a statistically significant difference.
Results
The three pungent ginger components inhibit CYP450 activities in vitro
The parameters used to determine the activities of the P450 enzymes are summarized in Table 1. A solvent control (methanol) was included in the assays; 1% methanol showed no detectable inhibitory effect against Vivid green, red or blue metabolism by CYP3A4 but had minimal inhibitory activity against Vivid blue metabolism by CYP2C19 (Table 2). However, 1% methanol inhibited Vivid CYP2C9 red metabolism by up to 40.6% and CYP2D6 blue metabolism by 27.8%. When the final concentration of methanol was reduced to 0.5% or 0.1%, the inhibition of P450s activities was either negligible or within an acceptable range (Table 2).
All three gingerols potently inhibited CYP2C9 activity, had moderate inhibitory effects on CYP2C19 and CYP3A4 activities, and had weak inhibitory effects on CYP2D6 activity (Table 2). Among the three compounds, 8-gingerol had the greatest inhibition against all four P450 enzymes, with IC50 values of 6.8 μmol/L for CYP2C9, 12.5 μmol/L for CYP2C19, 8.7 μmol/L for CYP3A4, and 42.7 μmol/L for CYP2D6.
Gingerol inhibition of CYP3A4 has no substrate-selective difference
Because of the potential influence of substrate selection on evaluating the effects of gingerols on the catalytic activity of CYP3A4, three fluorescent probes, Vivid DBOMF CYP3A4 green, Vivid BOMR CYP3A4 red and Vivid BOMCC CYP3A4 blue (Figure 1B), were used. The assay conditions are summarized in Table 1. The solvent effects of methanol (1%) on CYP3A4 green, red and blue metabolisms were negligible. For all the tested gingerols, the inhibition profiles determined with the green, red and blue substrates were essentially the same. No significant difference in the inhibitory potency against the three substrates was found for the same gingerol (Table 2). All three compounds inhibited CYP3A4 in a dose-dependent manner, with the potency decreasing in the following order: 8-gingerol > 10-gingerol > 6-gingerol (Figure 2A–2C).

Effects of the gingerols on the Vivid CYP3A4 green assay (A), Vivid CYP3A4 red assay (B) and Vivid CYP3A4 blue assay (C). Mean±SD of a percentage ratio of CYP3A4 reaction velocities to the velocity with no inhibitor present.
8- and 10-gingerol inhibit but 6-gingerol induces CYP3A4 mRNA expression in HepG2 cells
The effects of the three gingerols on CYP3A4 mRNA expression were different. RIF (the positive control) induced CYP3A4 expression by 138%±29%. In HepG2 cells, 8- and 10-gingerol decreased the expression of CYP3A4 by 58%±7% and 61%±4%, respectively, but 6-gingerol increased the CYP3A4 mRNA level by 119%±31% (Figure 3).

Effects of the gingerols on CYP3A4 mRNA expression in HepG2 cells. After normalization to β-actin levels, the results from three independent assays were averaged and the relative expression levels compared to the vehicle control were reported (mean±SD). RIF, Rifampicin. Differences were significant when compared to the vehicle control group (bP<0.05).
Discussion
Although microsomal assays are commonly used to evaluate the activity of P450 enzymes, fluorescent methods for determining P450 activity allow for high-throughput screening and more reproducible results21,22,23,24,25. Bell et al found that nearly 75% of marketed drugs yielded acceptable correlations between fluorogenic and HLM+LC-MS/MS assays for CYPs 3A4, 2D6, and 2C9, which supports the use of fluorogenic assays for rapid early risk binning based on IC50 values26. In the present study, fluorescent probes were used to study the effects of three pungent constituents of ginger on P450-mediated drug metabolism. The correlation between fluorescence assays and LC-MS/MS methods was confirmed in our preliminary study.
In the current study, 6-, 8-, and 10-gingerol substantially inhibited the four major P450 enzymes, with the exception that the inhibitory effects of gingerols against CYP2D6 were relatively weak. 6-, 8-, and 10-gingerol all exhibited strong inhibition against CYP2C9, with IC50 values of 12.1, 6.8, and 16.3 μmol/L, respectively. These values may have clinical implications. For example, 3.6±1.1 μmol/L of 6-gingerol, 0.84±0.47 μmol/L of 8-gingerol and 1.3±0.5 μmol/L of 10-gingerol were detected in human plasma 60 min after the ingestion of 2.0 g of ginger extract27. Compared to the corresponding IC50 values, a relatively high exposure of ginger extract may cause drug-diet interactions through P450 inhibition; such interactions may have an important implication in the clinical use of gingerols in combination with pharmacotherapy.
CYP2C9 represents approximately 20% of the hepatic CYP content in liver microsomes28. CYP2C9 is expressed at the highest level of the CYP2C family. Pharmacogenomic variations of CYP2C9 have been well documented. This isoform is involved in the metabolism of approximately 10% of all drugs, including nonsteroidal antiinflammatory drugs, oral antidiabetics, antiinfectives, hypnotics, antiepileptics, oral anticoagulants, such as warfarin, sulfonylureas, psychotropics, and angiotensin-2 antagonists29,30. It also metabolizes endogenous substrates, such as arachidonic acid and linolenic acid. CYP2C9 inhibition is involved in increases in the plasma concentration and toxicity of concomitant substrate drugs of CYP2C9, especially those with a narrow therapeutic index (eg, warfarin and phenytoin)31. Therefore, these factors should be taken into consideration when ginger is consumed.
The gingerols had significant inhibitory effects on CYP3A4. The CYP3A family contributes to approximately 50% of the total cytochrome P450 activity in the adult human liver and is involved in the metabolism of approximately 60% of therapeutic agents as well as the activation of toxic and carcinogenic substances32. CYP3A4 is the dominant CYP3A family enzyme expressed in the human liver and gastrointestinal tract33. Pharmacogenomic variations of CYP3A4 in humans have been indicated in many drug metabolism and drug-drug interactions. Because the effects on CYP3A4 might be substrate-dependent, as we found previously for the ginsenosides34, three fluorescent probes, Vivid CYP3A4 green, Vivid CYP3A4, red and Vivid CYP3A4 blue, were used to determine the effects of gingerols on CYP3A4. There was no significant difference in the inhibitory potency of these compounds.
In the present study, we also determined the effects of gingerols on CYP3A4 expression. The HepG2 cell line was employed in this study because it is a suitable surrogate of primary human hepatocytes to determine changes in CYP3A4 expression in the human body35. Based on the in vitro data, it appeared that 6-gingerol was a potent inhibitor of CYP3A4, and 8- and 10-gingerol also inhibited CYP3A4 transcription. Thus, attention should be paid when gingerols, especially 8- and 10-gingerol, are co-administered with other drugs, including herbal medicines that are metabolized by CYP3A4. 6-Gingerol potently induced CYP3A4 mRNA (similar to RIF) at a low concentration (10 μmol/L). At 10 μmol/L, 6-gingerol weakly inhibited CYP3A4 (approximately 20%). This finding indicates that 6-gingerol may mainly influence the transcription of CYP3A4 at a low concentration, which is usually the case in vivo. Thus, complex interactions between ginger and substrates of CYP3A4 can be deduced, and further studies are needed.
The result also suggested a structure-activity relationship (SAR) for the effects of gingerols on most of the P450 enzymes examined (Table 2). These gingerols are analogs and differ only in the length of the alkyl chain linked to the 4-hydroxy-3-methoxyphenyl scaffold (Figure 1A). 8-Gingerol, which has a moderate length of the aliphatic chain, showed the strongest inhibition against P450 enzymes. It appears that the moderate length of the alkyl chain allowed 8-gingerol to interact with P450s in an effective manner; however, further investigations are needed to confirm the molecular and structural basis of this finding.
In summary, our results indicate that the major pungent components of ginger can inhibit major human cytochrome P450 enzymes, which are responsible for the metabolism of most prescription drugs. Furthermore, the differential effects of gingerols on CYP3A4 gene expression may have complex consequences on the metabolism of different drugs. These findings highlight the importance of the safe use of ginger, especially in combination with clinically used drugs that utilize P450 enzymes as major metabolizing enzymes. This study may also serve as an example of drug-dietary supplement interactions in drug therapy.
Author contribution
Hui WANG directed the project, and along with Mian LI, defined the scope of the research; Mian LI and Pei-zhan CHEN designed methods and experiments and carried out the laboratory experiments; Mian LI analyzed the data, interpreted the results and drafted the manuscript; Qing-xi YUE and Jing-quan LI performed laboratory experiments and collected data; Rui-ai CHU and Wei ZHANG revised the manuscript; and Hui WANG reviewed the manuscript.
References
Kawai T, Kinoshita K, Koyama K, Takahashi K . Anti-emetic principles of Magnolia obovata bark and Zingiber officinale rhizome. Planta Med 1994; 60: 17–20.
Chrubasik S, Pittler MH, Roufogalis BD . Zingiberis rhizoma: a comprehensive review on the ginger effect and efficacy profiles. Phytomedicine 2005; 12: 684–701.
Grant KL, Lutz RB . Alternative therapies: ginger. Am J Health Syst Pharm 2000; 57: 945–7.
Wei W, Chang YL, Xiao DW, Ping L, Lian WQ . Simultaneous determination of 6-gingerol, 8-gingerol, 10-gingerol and 6-shogaol in rat plasma by liquid chromatography-mass spectrometry: Application to pharmacokinetics. J Chromatogr B Analyt Technol Biomed Life Sci 2009; 877: 671–9.
Govindarajan VS . Ginger — chemistry, technology, and quality evaluation: part 1. Crit Rev Food Sci Nutr 1982; 17: 1–96.
Young HY, Luo YL, Cheng HY, Hsieh WC, Liao JC, Peng WH . Analgesic and anti-inflammatory activities of 6-Gingerol. J Ethnopharmacol 2005; 96: 207–10.
Kiuchi F, Iwakami S, Shibuya M, Hanaoka F, Sankawa U . Inhibition of prostaglandin and leukotriene biosynthesis by gingerols and diarylheptanoids. Chem Pharm Bull (Tokyo) 1992; 40: 387–91.
Shoji N, Iwasa A, Takemoto T, Ishida Y, Ohizumi Y . Cardiotonic principles of ginger (Zingiber officinale Roscoe). J Pharm Sci 1982; 71: 1174–5.
Ueki S, Miyoshi M, Shido O, Hasegawa J, Watanabe T . Systemic administration of 6-Gingerol, a pungent constituent of ginger, induces hypothermia in rats via an inhibitory effect on metabolic rate. Eur J Pharmacol 2008; 584: 87–92.
Park KK, Chun KS, Lee JM, Lee SS, Surh YJ . Inhibitory effects of 6-Gingerol, a major pungent principle of ginger, on phorbol ester-induced inflammation, epidermal ornithine decarboxylase activity and skin tumor promotion in ICR mice. Cancer Lett 1998; 129: 139–44.
Park YJ, Wen J, Bang S, Park SW, Song SY . 6-Gingerol induces cell cycle arrest and cell death of mutant p53-expressing pancreatic cancer cells. Yonsei Med J 2006; 47: 688–97.
Ishiguro K, Ando T, Maeda O, Ohmiya N, Niwa Y, Kadomatsu K, et al. Ginger ingredients reduce viability of gastric cancer cells via distinct mechanisms. Biochem Biophys Res Commun 2007; 362: 218–23.
Lee SH, Cekanova M, Baek SJ . Multiple mechanisms are involved in 6-gingerol-induced cell growth arrest and apoptosis in human colorectal cancer cells. Mol Carcinog 2008; 47: 197–208.
Lee HS, Seo EY, Kang NE, Kim WK . 6-Gingerol inhibits metastasis of MDA-MB-231 human breast cancer cells. J Nutr Biochem 2008; 19: 313–9.
Haniadka R, Rajeev AG, Palatty PL, Arora R, Baliga MS . Zingiber officinale (ginger) as an anti-emetic in cancer chemotherapy: a review. J Altern Complement Med 2012; 18: 440–4.
Schwertner HA, Rios DC . High-performance liquid chromatographic analysis of 6-gingerol, 8-gingerol, 10-gingerol, and 6-shogaol in ginger-containing dietary supplements, spices, teas, and beverages. J Chromatogr B Analyt Technol Biomed Life Sci 2007; 856: 41–7.
Zick SM, Djuric Z, Ruffin MT, Litzinger AJ, Normolle DP, Alrawi S, et al. Pharmacokinetics of 6-gingerol, 8-gingerol, 10-gingerol, and 6-shogaol and conjugate metabolites in healthy human subjects. Cancer Epidemiol Biomarkers Prev 2008; 17: 1930–6.
Sparreboom A, Cox MC, Acharya MR, Figg WD . Herbal remedies in the United States: potential adverse interactions with anticancer agents. J Clin Oncol 2004; 22: 2489–503.
Moon YJ, Wang X, Morris ME . Dietary flavonoids: effects on xenobiotic and carcinogen metabolism. Toxicol In Vitro 2006; 20: 187–210.
Eric Y, Kathy A . Interaction of herbal constituents with cytochrome P450 enzymes. Alternative and Complementary Therapies 2007; 13: 239–47.
Hao M, Ba Q, Yin J, Li J, Zhao Y, Wang H . Deglycosylated ginsenosides are more potent inducers of CYP1A1, CYP1A2 and CYP3A4 expression in HepG2 cells than glycosylated ginsenosides. Drug Metab Pharmacokinet 2011; 26: 201–5.
Trubetskoy OV, Gibson JR, Marks BD . Highly miniaturized formats for in vitro drug metabolism assays using vivid fluorescent substrates and recombinant human cytochrome P450 enzymes. J Biomol Screen 2005; 10: 56–66.
Marks BD, Thompson DV, Goossens TA, Trubetskoy OV . Highthroughput screening assays for the assessment of CYP2C9*1, CYP2C9*2, and CYP2C9*3 metabolism using fluorogenic Vivid substrates. J Biomol Screen 2004; 9: 439–49.
Marks BD, Smith RW, Braun HA, Goossens TA, Christenson M, Ozers MS, et al. A high throughput screening assay to screen for CYP2E1 metabolism and inhibition using a fluorogenic vivid p450 substrate. Assay Drug Dev Technol 2002; 1: 73–81.
Marks BD, Goossens TA, Braun HA, Ozers MS, Smith RW, Lebakken C, et al. High-throughput screening assays for CYP2B6 metabolism and inhibition using fluorogenic vivid substrates. AAPS PharmSci 2003; 5: E18.
Bell L, Bickford S, Nguyen PH, Wang J, He T, Zhang B, et al. Evaluation of fluorescence- and mass spectrometry-based CYP inhibition assays for use in drug discovery. J Biomol Screen 2008; 13: 343–53.
Zick SM, Ruffin MT, Djuric Z, Normolle D, Brenner DE . Quantitation of 6-, 8- and 10-gingerols and 6-shogaol in human plasma by high-performance liquid chromatography with electrochemical detection. Int J Biomed Sci 2010; 6: 233–40.
Nassar AF, Hollenberg PF, Scatina JA . Drug metabolism handbook: concepts and applications. New Jersey: John Wiley & Sons, Inc; 2009.
Cohen LH, Remley MJ, Raunig D, Vaz AD . In vitro drug interactions of cytochrome p450: an evaluation of fluorogenic to conventional substrates. Drug Metab Dispos 2003; 31: 1005–15.
Magnus IS, Sarah CS, Alvin G, Cristina RA . Influence of cytochrome P450 polymorphisms on drug therapies: Pharmacogenetic, pharmacoepigenetic and clinical aspects. Pharmacol Ther 2007; 116: 496–526.
Miners JO, Birkett DJ . Cytochrome P450 2C9: an enzyme of major importance in human drug metabolism. Br J Clin Pharmacol 1998; 45: 525–38.
Cascorbi I . Genetic basis of toxic reactions to drugs and chemicals. Toxicol Lett 2006; 162: 16–28.
Lamba JK, Lin YS, Schuetz EG, Thummel KE . Genetic contribution to variable human CYP3A-mediated metabolism. Adv Drug Deliv Rev 2002; 54: 1271–94.
Hao M, Zhao Y, Chen P, Huang H, Liu H, Jiang H, et al. Structure-activity relationship and substrate-dependent phenomena in effects of ginsenosides on activities of drug-metabolizing P450 enzymes. PLoS One 2008; 3: e2697.
Westerink WM, Schoonen WG . Cytochrome P450 enzyme levels in HepG2 cells and cryopreserved primary human hepatocytes and their induction in HepG2 cells. Toxicol In Vitro 2007; 21: 1581–91.
Acknowledgements
We thank Ms Yuan-yuan DAI at the Cancer Hospital/Institute, Chinese Academy of Medical Sciences, for her generous gift of sulfaphenazole. This study was supported by grants from the One Hundred Talents Program and Knowledge Innovation Program (KSCX2-EW-R-10) of the Chinese Academy of Sciences, the National Natural Science Foundation (81125020, 91029715, 31070680, 31101261, 81242002, and 31200569), the Ministry of Science and Technology of China (2012BAK01B00, 2011BAK10B00, and 2009CB919000), the Science and Technology Commission of Shanghai Municipality (12XD1407000, 12431900500, and 10391902100), and Xuhui Central Hospital (CRC2011001 and CRC2011004), Director Foundation (20090101) and the Food Safety Research Center and Key Laboratory of Nutrition and Metabolism of INS, SIBS, CAS. Pei-zhan CHEN and Qing-xi YUE were partially supported by the SA-SIBS scholarship program.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Li, M., Chen, Pz., Yue, Qx. et al. Pungent ginger components modulates human cytochrome P450 enzymes in vitro. Acta Pharmacol Sin 34, 1237–1242 (2013). https://doi.org/10.1038/aps.2013.49
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2013.49
Keywords
This article is cited by
-
Herb-anticancer drug interactions in real life based on VigiBase, the WHO global database
Scientific Reports (2022)
-
Dual protective effect of ginger and rosemary extracts against CCl4-induced hepatotoxicity in rats
Environmental Science and Pollution Research (2018)
