
Abstract
Lipid rafts have been known to be platforms to initiate cellular signal transduction of insulin-like growth factor (IGF) inducing skeletal muscle differentiation and hypertrophy. Here, tripartite motif 72 (TRIM72), with a really interesting new gene (RING)-finger domain, a B-box, two coiled-coil domains, and a SPRY (SPla and RYanodine receptor) domain, was revealed to be predominantly expressed in the sarcolemma lipid rafts of skeletal and cardiac muscles. Adenoviral TRIM72 overexpression prevented but RNAi-mediated TRIM72 silencing enhanced C2C12 myogenesis by modulating the IGF-induced insulin receptor substrate-1 (IRS-1) activation through the molecular association of TRIM72 with IRS-1. Furthermore, myogenic activity was highly enhanced with increased IGF-induced Akt activation in the satellite cells of TRIM72−/− mice, compared to those of TRIM72+/+ mice. Because TRIM72 promoter analysis shows that two proximal E-boxes in TRIM72 promoter were essential for MyoD- and Akt-dependent TRIM72 transcription, we can conclude that TRIM72 is a novel antagonist of IRS-1, and is essential as a negative regulator of IGF-induced muscle differentiation.
Similar content being viewed by others


Main
To execute rapid and efficient cellular signaling, there should be specific membrane platforms that organize receptors and their downstream signaling molecules. The platforms, so-called lipid rafts, are mainly composed of cholesterol and glycosphingolipids.1, 2 Cholesterol and saturated acyl chains of glycosphingolipids are tightly packed together, forming lipid rafts among ‘the sea of phospholipids’ in the plasma membrane. Thus, lipid rafts have been isolated from other cellular organelles by using their detergent insolubility and low density in sucrose gradient ultracentrifugation.3 The lipid rafts also generate flask-shaped plasma membrane invaginations, called caveolae, with the help of caveolin (Cav) and cavin proteins.4, 5 Thus, Cav-1 or -3 has been used as a lipid raft or caveoli marker protein. Lipid rafts and caveolae have been known to be involved in different cellular events such as cellular differentiation, immune response, and cell migration.6, 7, 8
To find novel lipid raft proteins, and gain new insights for the function of lipid rafts, we along with other groups have identified lipid raft proteins by proteomic analysis.8, 9, 10, 11, 12 Previously, we identified a novel lipid raft protein, a receptor of globular C1q (gC1qR), by the differential proteomic analysis of lipid rafts from 3T3 L1 preadipocytes and adipocytes, and showed that gC1qR is an essential signaling molecule involved in adipogenesis and insulin signal transduction.8 This result shows that the differential proteomic analyses of lipid rafts are very powerful tools to find novel signaling molecules involved in cellular differentiation, such as adipogenesis, neurogenesis, and myogenesis.
Insulin-like growth factors, IGF-I and IGF-II, are essential for skeletal muscle development, hypertrophy, and regeneration.13, 14 Skeletal precursor stem cells, satellite cells, lose their muscle differentiation activity when treated with anti-IGF-II antibody or anti-sense IGF-II,15, 16 whereas IGF-I overexpression in mice leads to the increased muscle mass and force generation as well as significant hypertrophy and hyperplasia at all ages.17 IGF-I and IGF-II transduce cellular signaling through the IGF-I receptor (IGFR) that subsequently recruits insulin receptor substrate-1 (IRS-1). The recruited IRS-1 activates phosphoinositide 3-kinase (PI(3)K)/Akt/mTOR and Ras/Raf/MEK/ERK pathways. The Ras/Raf/MEK/ERK pathway controls muscle fiber type whereas the PI(3)K/Akt pathway induces muscle differentiation and hypertrophy.18, 19 The activated Akt targets a mammalian target of rapamycin (mTOR), glycogen synthase kinase 3β (GSK3β), and forkhead box O (FOXO). The phosphorylated GSK3β and FOXO lead to stimulate muscle hypertrophy and to suppress muscle atrophy, respectively.20, 21, 22, 23 However, mTOR is not involved in IGF-dependent muscle differentiation because muscle differentiation is not affected by the mTOR inhibitor rapamycin in the presence of IGF.15 Lipid rafts might have a crucial role in skeletal muscle differentiation and hypertrophy because lipid rafts enrich IGF signaling molecules such as IGFR, IRS-1, PI(3)K, Akt, and GSK3β.24, 25, 26, 27 Thus, novel signaling molecules involved in skeletal muscle differentiation could be disclosed by the proteomic analysis of lipid rafts.
Several tripartite motif (TRIM)-containing proteins, which are composed of a really interesting new gene (RING) finger, one or two B-boxes, and two coiled-coil (CC) domains,28 are expressed in skeletal muscle to regulate muscle coordination, atrophy, and repair. TRIM54, also called muscle-specific RING finger-1 (MURF-1), is upregulated in skeletal muscle under atrophic conditions to ubiquitinate and degrade myofibrillar proteins such as titin and nebulin.20, 29 Indeed, atrophy-induced loss of muscle mass is prevented in TRIM54/MURF-1-disrupted mice.20 TRIM55/MURF-2 is also associated with myofibril components (titin) and microtubules.30 The TRIM55/MURF-2 has been determined to be necessary for the structure of stable microtubules and intermediate filaments because its knockdown in skeletal myoblasts prevents myoblast fusion and myofibrillogenesis. TRIM32 and TRIM41/RINCK are also highly expressed in skeletal muscle and are involved in the ubiquitination of actin and protein kinase C βII, respectively.31, 32 Another TRIM family protein TRIM72/MG53 is specifically expressed in the plasma membrane of skeletal muscle, and it has a critical role in membrane repair after acute muscle injury.33 However, the molecular mechanism of TRIM72/MG53 has never been investigated in skeletal muscle differentiation.
Here, we identified a lipid raft protein, TRIM72, by the differential two-dimensional electrophoresis (2-DE) of lipid rafts from C2C12 myoblasts and myotubes. We have investigated the role of TRIM72 in myogenesis after overexpressing and downregulating TRIM72 by using adenoviral system and RNAi silencing. Myogenesis and IGF-I-induced IRS-1 tyrosine phosphorylation were prevented by TRIM72 overexpression but were enhanced by TRIM72 knockdown. Moreover, TRIM72 promoter activity was dependent on MyoD and Akt. Thus, we propose that TRIM72 is a novel negative feedback regulator of IGF-induced myogenesis.
Results
TRIM72 is specifically expressed in the sarcolemma lipid rafts of skeletal muscle and heart
We isolated detergent-resistant lipid rafts from C2C12 myoblasts and myotubes based on the Triton X-100 insolubility and low density (Supplementary Figure S1A); C2C12 mononucleated myoblasts were differentiated into multinucleated myotubes by incubating them with 2% horse serum at confluence for 2 days. The lipid raft proteins were resolved by 2-DE and visualized by silver staining. Myotube-specific lipid raft proteins indicated by circles in 2-DE gel profiles were identified by liquid chromatography-electrospray ionization-tandem mass spectrometry (LC-ESI-MS/MS) (Supplementary Figure S1B). Among these spots, four protein spots with a molecular weight of 55 kDa (Figure 1a) were identified as TRIM72 (Supplementary Figure S1C). The TRIM72 gene contains TRIM/RBCC (tripartite motif or N-terminal RING finger/B-box/coiled-coil) and SPRY (SP la and RYanodine receptor) domains, which are necessary for ubiquitin E3 ligase activity and protein–protein interaction, respectively34 (Figure 1b and Supplementary Figure S1D).

Identification of TRIM72 in the lipid rafts of skeletal muscle. (a) C2C12 myotube-specific TRIM72 spots were identified by the comparative 2-DE analysis of lipid raft proteins isolated from myoblasts and myotubes; C2C12 myoblasts were differentiated into myotubes for 2 days. Areas containing red circles in Supplementary Figure S1B are enlarged. TRIM72 spots are indicated by dotted circles. (b) Gene analysis of TRIM72 shows a RING finger domain, a B-box, two coiled-coil domains, and a SPRY domain. (c) Northern blotting analysis of TRIM72 in various mouse organs. (d) Western blotting of TRIM72 and caveolin-3 (Cav-3) in various organs obtained from 12-week-old male mice. (e) Reverse transcription-polymerase chain reaction (RT-PCR) analysis of TRIM72, myogenin, MyoD, Cav-3, and myosin heavy chain (MyHC) during C2C12 myogenesis by using actin as a loading control. C2C12 myoblasts were differentiated to myotubes for the indicated times. (f) Western blotting of TRIM72, Cav-3, myogenin, MyHC, MyoD, and Myf5 during C2C12 myogenesis. Ponceau S staining was used for a loading control. (g) Immunofluorescence analysis of TRIM72 and Cav-3 in mouse gastrocnemius skeletal muscle obtained from 12-week-old male mice. (h) Enrichment of TRIM72 and Cav-3 in detergent-resistant lipid rafts. Detergent-resistant lipid rafts were isolated from 4-days-differentiated C2C12 myotubes and mouse gastrocnemius muscle on the basis of detergent insolubility and low density, fractionated after a discontinuous sucrose gradient, and analyzed by western blotting of TRIM72 and Cav-3. (i) TRIM72 was postnatally expressed in mouse skeletal muscle and heart. Mouse skeletal muscle and heart were obtained at embryonic (E) and postnatal (P) stages and analyzed by western blotting
TRIM72 mRNA and its protein products were exclusively expressed in mouse skeletal muscle and heart (Figure 1c and d). TRIM72 began to be expressed with myogenesis markers such as myogenin, Cav-3, and myosin heavy chain (MyHC) at the first day of C2C12 myogenesis, as determined by reverse transcription-polymerase chain reaction (RT-PCR) and western blotting (Figure 1e and f). As expected, TRIM72 was colocalized with Cav-3 in C2C12 myotubes and mouse skeletal muscle and heart (Figure 1g and Supplementary Figure S1E), and was enriched in the lipid rafts (Figure 1h), indicating that TRIM72 is indeed a novel lipid raft protein in skeletal muscle and heart. Interestingly, TRIM72 was highly expressed in the skeletal muscle and heart isolated from the mice of postnatal stage (Figure 1i).
TRIM72 is a negative regulator of myogenesis
To understand the role of TRIM72 in myogenesis, we treated C2C12 myoblasts with an adenoviral vector with TRIM72 (Ad-TRIM72) or small-interference RNA (siRNA) for TRIM72 (si-TRIM72) and differentiated to myotubes for 4 days. As shown in Figure 2c, TRIM72 was highly overexpressed by using Ad-TRIM72 and strongly downregulated by si-TRIM72 oligomer. Myogenesis was monitored by H&E staining, MyHC immunofluorescence, western blotting of myogenic marker proteins (MyHC and Cav-3), and myogenic index. As shown in Figure 2a–d, myogenesis was completely prevented by TRIM72 overexpression, but enhanced by TRIM72 knockdown. To make sure the effect of TRIM72 knockdown on the enhancement of myogenesis, we treated C2C12 myoblasts with four different si-TRIM72 oligomers and differentiated to myotubes. Myogenesis was also monitored by H&E staining and western blotting of myogenic marker proteins. As shown in Supplementary Figure S2, all si-TRIM72 oligomers enhanced C2C12 myogenesis. Because Cai et al. show that TRIM72 knockdown abolishes C2C12 myogenesis, we determined the effect of TRIM72 on C2C12 myogenesis by using the si-TRIM72/MG53 oligomer with the same sequence used by Cai et al. As shown in Supplementary Figure S2E, TRIM72 expression level was significantly decreased in C2C12 myotubes treated with the si-TRIM72/MG53 oligomer, indicating that the si-TRIM72/MG53 oligomer is also an excellent siRNA for TRIM72 knockdown. As C2C12 myogenesis was monitored by H&E staining, myogenic index, and western blotting with anti-Cav-3, MyHC, and myogenin, the si-TRIM72/MG53 oligomer was determined to enhance myogenesis as shown in Supplementary Figure S2C–E. Taken together, with these data shown in Figure 2 and Supplementary Figure S2, we can conclude that TRIM72 is a negative regulator of myogenesis.

TRIM72 is a myogenesis inhibitor. C2C12 myoblasts were treated with Ad-TRIM72 (2 × 1010 viral particles per ml) or si-TRIM72 (200 nM) for 1 day and differentiated to myotubes for 4 days. Skeletal muscle differentiation was monitored by H&E staining (a), MyHC immunofluorescence, and DAPI staining (b), western blotting of TRIM72, Cav-3, and MyHC, using Ponceau S staining as a loading control (c), and myogenic index (d). Myogenic index was determined as the ratio of the nuclear number in myotubes (C2C12 cells with two or more nuclei) to the total nuclear number in H&E-stained fields. *P<0.01. Ad-EV, adenovirus with an empty vector; Ad-TRIM72, adenovirus with a mouse TRIM72 gene; Si-control, siRNA with a scrambled sequence; Si-TRIM72, TRIM72-specific siRNA
To further test the role of TRIM72 on human skeletal muscle, we differentiated human primary skeletal muscle cells to myotubes for the indicated times. As shown in Supplementary Figure 3A and B, TRIM72 mRNA and its protein product were gradually increased with myogenic markers in differentiating human primary muscle cells. When human primary skeletal muscle cells were treated with Ad-TRIM72 and differentiated to human myotubes for 4 days, myogenesis was prevented by TRIM72 overexpression as determined by MyHC immunofluorescence, western blotting of myogenic marker proteins, and the nuclear number in MyHC-stained myotubes (Supplementary Figure 3C–E). Because siRNA was not efficiently transfected into human primary muscle cells, we could not investigate the effect of TRIM72 knockdown on the myogenesis of human muscle cells. Taken together all these data, we can conclude that TRIM72 is a negative regulator in skeletal muscle differentiation.
TRIM72 inhibits IGF-induced IRS-1 activation
Because IGF is a major growth factor regulating skeletal muscle differentiation and hypertrophy,13, 14 we tested the possible involvement of TRIM72 in the IGF-I-mediated activation of the IGFR/IRS-1/PI(3)K/Akt signaling cascade. IGF-I-induced Akt phosphorylation was inhibited in myoblasts by TRIM72 overexpression, but was enhanced in myotubes by TRIM72 knockdown (Figure 3a), showing that TRIM72 prevented IGF-I-induced Akt activation. In contrast, IGF-I-induced Erk1/2 phosphorylation was not changed by TRIM72 overexpression or knockdown, suggesting that the IGFR/Raf/MAPK pathway might not be affected by TRIM72. The TRIM72 inhibition of IGF-I-induced Akt activation prompted us to test the effect of TRIM72 on PI(3)K activation. Figure 3b shows that IGF-I-induced PI(3)K activation was abrogated in myoblasts with TRIM72 overexpression, but was enhanced in myotubes with TRIM72 knockdown (Figure 3b). Next, we tested the TRIM72 effect on the IGF-I-induced tyrosine phosphorylation of IGFR and IRS-1 (Figure 3c). The tyrosine phosphorylation of IGFR was not changed by TRIM72 overexpression or knockdown in both myoblasts and myotubes exposed to IGF-I. However, the IGF-I-induced tyrosine phosphorylation of IRS-1 was decreased by TRIM72 overexpression, but increased by TRIM72 knockdown. Moreover, the molecular association of IRS-1 with PI(3)K after IGF-I stimulation was weakened by TRIM72 overexpression, but fortified by TRIM72 knockdown (Figure 3c). Because TRIM72 inhibits IGF-I-induced IRS-1 activation, TRIM72 might negatively regulate IGF-I-induced glucose uptake in skeletal muscle. To address the issue, we determined glucose uptake in C2C12 myotubes treated with si-control or si-TRIM72 oligomer. As shown in Figure 3d, TRIM72 knockdown enhanced IGF-I-induced glucose uptake in myotubes.

TRIM72 suppresses IGF-induced IRS-1 activation. (a) TRIM72 was overexpressed with Ad-TRIM72 in C2C12 myoblasts for 48 h or knockdowned with si-TRIM72 in C2C12 myoblasts that were further differentiated to myotubes for 4 days. TRIM72-overexpressing myoblasts and TRIM72-knockdowned myotubes were stimulated by IGF-I (100 ng/ml) for the indicated times. TRIM72, pAkt, total Akt, pErk1/2, and total Erk1/2 were analyzed by western blotting, using actin as a loading control. (b) IGF-dependent PI(3)K activity was measured in mouse TRIM72-overexpressing myoblasts and TRIM72-knockdowned myotubes. (c) After treating TRIM72-overexpressing myoblasts and TRIM72-knockdowned myotubes with IGF-I (100 ng/ml) for 0, 2, and 5 min, the tyrosine phosphorylation of IGFR and IRS-1, and the molecular association of IRS-1 with PI(3)K were investigated by endogenous immunoprecipitation. IP, immunoprecipitation; WB, western blotting. (d) C2C12 myoblasts were treated with si-control or si-TRIM72 for 1 day and differentiated to myotubes for 3 days. The C2C12 myotubes were incubated with 3H-deoxyglucose (10 μCi/ml) and IGF-I (200 ng/ml) for 30 min. Cell-associated radioactivity was measured and normalized to protein concentration. Each panel is the representative result of three separate experiments. The expression level of TRIM72 was determined by western blotting (inset panel)
TRIM72 is associated with IRS-1
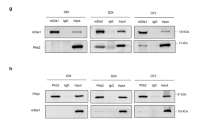
The TRIM72 inhibition of IRS-1 activation indicates that TRIM72 could target IRS-1. To explore the possibility, we tested the molecular association of TRIM72 with IRS-1 by endogenous immunoprecipitation using C2C12 myotubes stimulated with IGF-I. TRIM72 and IRS-1 were tightly associated together even in the absence of IGF-I stimulation (Figure 4a). To analyze the binding domains of TRIM72 and IRS-1, we transiently expressed various deletion mutants of both genes (Figure 4b) in HEK-293 cells, and the binding activity was investigated by co-immunoprecipitation. As shown in Figure 4c and d, the middle and C-terminal regions of human IRS-1 (hIRS-1) and the CC domain of human TRIM72 (hTRIM72) were required for the molecular interaction because CC domain-deleted hTRIM72 mutants (SPRY and ΔCC) did not bind to hIRS-1.

(a) Molecular interaction of TRIM72 with IRS-1 was determined by endogenous immunoprecipitation in the myotubes stimulated with IGF-I (100 ng/ml) for the indicated times. (b) Deletion constructs for human IRS-1 (hIRS-1) and human TRIM72 (hTRIM72). Full, full-length; PH, pleckstrin homology domain; PTB, phosphotyrosine binding domain; C-term, C-terminal region. (c and d) 293 cells were co-transfected with Myc-tagged hTRIM72 (2 μg) and various Flag-tagged hIRS-1 deletion mutants (6 μg) (c) or with Flag-tagged hIRS-1 (6 μg) and Myc-tagged hTRIM72 deletion mutants (2 μg) (d) for 24 h. After 12 h of MG132 (5 μM) treatment, molecular interaction was determined by immunoprecipitation
To further confirm the involvement of IRS-1 in the enhanced muscle differentiation by TRIM72 knockdown, we treated C2C12 myoblasts with si-control alone, si-TRIM72 alone, si-IRS-1 alone, or both si-TRIM72 and si-IRS-1 and further differentiated to myotubes for 3 days. As determined by H&E staining and western blotting of myogenesis marker proteins (Cav-3 and MyHC) (Supplementary Figure S4), myogenesis was prevented by IRS-1 knockdown. Moreover, the enhanced myotube formation by TRIM72 knockdown was greatly decreased by IRS-1 depletion, indicating that IRS-1 is a downstream signaling molecule of TRIM72 that is required for the negative regulation of myogenesis.
TRIM72 disruption enhances the myogenesis of satellite cells
Next, we monitored the expression level of TRIM72 during the myogenesis of mouse satellite cells that are skeletal muscle stem cells. Satellite cells were isolated from the hind limb muscles of 10-week-old male wild-type mice and differentiated to myotubes. The mRNA and protein level of TRIM72 were increased with other myogenic markers such as MyHC, myogenin, and Cav-3 during myogenesis (Supplementary Figure S5). To confirm the negative effect of TRIM72 on myogenesis, we generated TRIM72-disrupted mice as shown in Figure 5a. TRIM72 disruption was confirmed by RT-PCR and western blotting (Figure 5b and c). TRIM72−/− mice were viable, and their skeletal muscle showed no muscle pathologies until 17 weeks because central nuclei were rarely found in DAPI-stained muscle fibers (unpublished data). Satellite cells were obtained from TRIM72+/+ and TRIM72−/− mice and monitored for their myogenic capacities by MyHC immunofluorescence; western blotting for TRIM72, MyHC, myogenin, and Cav-3; and myogenic index (Figure 5d–f). Myogenesis was highly enhanced in the satellite cells obtained from TRIM72−/− mice compared to those from wild-type mice, indicating that TRIM72 negatively regulates myogenesis. IGF-I-induced Akt phosphorylation was also monitored in the myotubes differentiated from wild-type and TRIM72−/− satellite cells. IGF-I-dependent Akt phosphorylation was increased in TRIM72−/− myotubes compared to wild-type myotubes (Figure 5g). In contrast, Erk1/2 was equally phosphorylated by IGF-I in both TRIM72+/+ and TRIM72−/− satellite myotubes, indicating that TRIM72 does not affect the IGF-I-induced Erk1/2 activation.

TRIM72 disruption enhances myogenesis in satellite cells. (a) Schematic representation for the generation of TRIM72-disrupted mice. Exons 1, 2, and 3 of the mouse TRIM72 gene were replaced by a cassette containing the neomycin resistance gene driven by the PGK promoter (PGK-neo). The targeting vector also possessed a positive selection marker (PGK-tk). Forward and inverted arrowheads indicate the primers used for genotyping and PCR. (b) PCR analysis of TRIM72 levels in skeletal muscle obtained from TRIM72+/+, TRIM72+/−, and TRIM72−/− mice. PCR products of wild-type and mutant TRIM72 were 817 and 2867 bp, respectively. (c) Skeletal muscles were isolated from TRIM72+/+, TRIM72+/−, and TRIM72−/− mice and analyzed by western blotting with an anti-TRIM72 antibody. Ponceau S staining was used as a loading control. (d–f) Satellite cells were obtained from 4-week-old female TRIM72+/+ and TRIM72−/− mice, and differentiated to myotubes for 36 h. Myogenesis was monitored by MyHC immunofluorescence and DAPI staining (d), western blotting for TRIM72, MyHC, myogenin, Cav-3, and actin (e), and myogenic index (f). Myogenic index was determined from the MyHC-stained cells. *P<0.01 (g) Myotubes were differentiated from TRIM72+/+ and TRIM72−/− satellite cells for 48 h, serum-starved for 4 h, and treated with IGF-I (200 ng/ml) for the indicated times. TRIM72, pAkt, total Akt, pErk1/2, and total Erk1/2 were analyzed by western blotting. Ponceau S staining was used as a loading control
TRIM72 transcription requires MyoD and Akt
Because TRIM72 is gradually increased during myogenesis (Figure 1e and f), it is tempting to speculate that the TRIM72 promoter contains E-boxes that are binding sites for the activated MyoD.35 Indeed, the TRIM72 promoter contained five E-boxes (Figure 6a). To determine the E-boxes required for TRIM72 expression, we transfected the luciferase reporters with various truncation mutants in TRIM72 promoter into C2C12 myoblasts that were consecutively differentiated to myotubes. After 24 h of myotube differentiation, luciferase activity was measured for the promoter activity. Both E1 and E2 boxes were required for TRIM72 transcription during myogenesis because TRIM72 promoter activity was dramatically decreased when both E1 and E2 boxes were truncated or mutated (Figure 6a). In different cell lines such as C2C12 myoblasts, HEK-293 cells, and NIH3T3 cells, TRIM72 promoter activity was significantly increased by the transient expression of MyoD, myogenin, and Mrf4 (Figure 6b, and Supplementary Figure S6A and B). Because the mRNA level of TRIM72 was significantly decreased with the reduced level of myogenin and Cav-3 by RNAi silencing of MyoD in C2C12 myotubes (Figure 6c), MyoD might be a master transcription factor for TRIM72. In addition, MyoD was specifically associated with the E1 and E2 boxes of TRIM72 promoter in myotubes but not in myoblasts, as determined by chromatin immunoprecipitation (ChIP) (Figure 6d), indicating that MyoD activated during myogenesis is required for TRIM72 transcription.

TRIM72 expression is regulated by MyoD through a PI(3)K/Akt pathway. (a) The promoter of TRIM72 contains five E-boxes. The firefly luciferase gene was fused to various mutants of the TRIM72 promoter. C2C12 myoblasts grown in 12-well plates were transfected with TRIM72 reporters (0.8 μg) and β-Gal (0.8 μg) for 24 h. After 48 h of myotube differentiation, luciferase activity was normalized by β-Gal activity. Relative luciferase activity was calculated by the ratio of normalized luciferase activity in each experimental group to that in control group (EV-transfected cells). (b) MyoD, Myf5, myogenin, or Mrf4 (each 0.6 μg) was co-transfected with 883 bp TRIM72 reporter (0.6 μg) and β-Gal (0.6 μg) for 24 h into C2C12 myoblasts grown in 12-well plates. After 48 h of transfection, the relative luciferase activity was determined. The expression level of MyoD, Myf5, myogenin, and Mrf4 was determined by western blotting (inset panel). (c) RNAi silencing of MyoD abolishes the transcriptional activation of TRIM72. C2C12 myoblasts were treated with si-MyoD (200 nM) for 24 h, and differentiated to myotubes for 24 h. The mRNA level of MyoD, myogenin, TRIM72, Cav-3, and GAPDH was determined by RT-PCR. (d) MyoD binds to TRIM72 promoter. The chromatin was harvested from C2C12 myoblasts and 2-day-differentiated myotubes, and immunoprecipitated with anti-MyoD antibody. The precipitated chromatin was analyzed by PCR for TRIM72 promoter. ‘B’ indicates myoblasts and ‘T’ myotubes. (e) TRIM72 requires PI(3)K activation for its transcription. The 883 bp TRIM72 reporter (20 μg) was transfected for 24 h into C2C12 myoblasts grown in a 100-mm dish. The cells were transferred to 12-well plates, further grown for 24 h, and treated with IGF-I (100 ng/ml) in the presence of LY294002 (10 μM), rapamycin (100 nM), LiCl (5 mM), or PD98059 (10 μM). After 24 h of IGF-I treatment, relative luciferase activity was calculated by the ratio of luciferase activity in drug-treated cells to that in control cells. (f) The 883 bp TRIM72 promoter was co-transfected with 1 μg of constitutively active Akt, FOXO1, or GSK3β (CA-Akt, CA-FOXO1, or CA-GSK3β) with 0.8 μg of β-gal for 24 h in myoblasts grown in 12-well plates. After 24 h of differentiation, the relative luciferase activity was determined. The expression level of CA-Akt, CA-FOXO1, and CA-GSK3β was determined by western blotting (inset panel). (g) The 883 bp TRIM72 promoter was co-transfected with si-control or si-Akt with β-gal (0.8 μg) for 24 h in C2C12 myoblasts grown in 12-well plates. After 24 h of differentiation, the relative luciferase activity was determined. The expression level of Akt and actin was determined by western blotting (inset panel). (h) The expression level of TRIM72 is highly increased by CA-Akt overexpression. C2C12 myoblasts were treated with empty vector (EV) or Myc-tagged CA-Akt (Myc-CA-Akt, 4 μg in six-well plate) for 24 h, and differentiated to myotubes for further 24 h. The expression level of TRIM72, myogenin, Akt, and GSK3β was determined by western blotting
Next, we dissected the IGF-I signaling pathway inducing TRIM72 transcription by pharmacological approaches. TRIM72 promoter activity in IGF-I-dependent C2C12 myotube differentiation was abolished by inhibiting PI(3)K with LY294002 but not dramatically changed by preventing mTOR with rapamycin or GSK3β with LiCl (Figure 6e). In addition, MyoD-dependent TRIM72 promoter activity in C2C12 myoblasts was also dramatically decreased by inhibiting PI(3)K but not affected by preventing mTOR or GSK3β (Supplementary Figure S6C). The TRIM72 promoter activity was increased 2.5-fold by the transient expression of constitutively active Akt (CA-Akt), but not changed by that of CA-FOXO1 or CA-GSK3β in C2C12 myotubes (Figure 6f). In addition, the TRIM72 promoter activity was significantly decreased by si-Akt (Figure 6g). Because TRIM72 transcription was dependent on Akt activation, the expression level of TRIM72 protein might be increased in myotubes overexpressing CA-Akt. To test the possibility, we overexpressed CA-Akt in myoblasts that were further differentiated to myotubes for 24 h, and monitored the expression level of TRIM72 by western blotting. As shown in Figure 6h, the expression level of TRIM72 was significantly increased by CA-Akt. This result indicates that TRIM72 expression is induced by Akt that is activated during myogenesis. With the pharmacological and genetic studies, we can conclude that TRIM72 transcription requires the MyoD activated by PI(3)K/Akt pathway, independent of mTOR, FOXO1, and GSK3β.
Discussion
There has been a long-standing question how a mammalian organ regulates its size. A right gastrocnemius skeletal muscle of mouse never outgrows to a left one, suggesting that there might be a ‘size checkpoint’ operating at the organs’ total mass. It is still mysterious, however, what kinds of mammalian genes operate as ‘size checkpoints’ to set limits on organ size. Accumulating evidences indicate that an IGF/IGFR/IRS/PI(3)K/Akt/mTOR/S6K signaling pathway is prerequisite to increasing mammalian organ size.36 Although PI(3)K, Akt, mTOR, and S6K have been shown to positively regulate cell size in skeletal muscle, negative regulators of IGF signaling such as PTEN and tuberous sclerosis complex 1 and 2 have never been reported to be upregulated or activated to limit the size of skeletal muscle.
It has been known that the transcriptional property of MyoD is controlled through PI(3)K/Akt signaling pathway because dominant negative Akt abolishes MyoD-dependent muscle differentiation.37 Akt mediates the formation of skeletal muscle transcriptosome, which is composed of MyoD, p300, and p300/CBP-associated factor through direct phosphorylation of p300.38 PI(3)K/Akt blockage prevents the association of p300 with MyoD and p300/CBP-associated factor, and the formation of active transcriptosome. The transcriptional activation of TRIM72 should be regulated by PI(3)K/Akt pathway because TRIM72 promoter contains two proximal E-boxes that are essential binding sites for MyoD. Indeed, our pharmacological and genetic analyses show that transcriptional activation of TRIM72 was dependent on PI(3)K/Akt pathway. Another myogenic transcription factor myogenin, which is highly expressed by the activated MyoD and binds to the E-boxes of myogenic target genes, such as Cav-3 and MyHC, was revealed to be another crucial transcription factor for TRIM72 expression. Indeed, TRIM72 transcriptional activity was highly increased by myogenin overexpression in C2C12 myotubes, NIH3T3 cells, and HEK-293 cells (Figure 6b, and Supplementary Figure S6A and B), and TRIM72 mRNA level was dramatically decreased by RNAi silencing of myogenin as well as that of MyoD during C2C12 myogenesis (unpublished data). Here, we propose a model in which TRIM72, was expressed by the IGF/IGFR/IRS-1/PI(3)K/Akt pathway, and targets IRS-1, where it interferes with IGF-I-induced IRS-1 phosphorylation. These actions suppress PI(3)K and Akt activation, which negatively regulates excess skeletal muscle differentiation and hypertrophy (Figure 7).

A schematic representation showing TRIM72 as a negative feedback regulator during myogenesis. This model shows that TRIM72 expression is mediated by IGF/IGFR/IRS-1/PI(3)K/Akt/MyoD signaling pathway. After its expression, TRIM72 interacts with and negatively regulates IRS-1
It has been reported that TRIM72 is necessary for sarcolemmal membrane repair after acute injury. TRIM72 is associated with intracellular vesicles that are recruited into sarcolemmal membrane in myofibers.33 Skeletal muscles of TRIM72−/− mice show defective membrane-repair capacity with progressive myopathy and reduced exercise capability after acute injury. Cai et al.39 also proposed that TRIM72 might be a positive regulator of myogenesis because TRIM72 knockdown prevents C2C12 myogenesis. However, TRIM72−/− young mice do not show any severe muscular abnormalities in the absence of injuries,33 and primary satellite cells from TRIM72−/− neonates are capable to be differentiated to myotubes (Figure 5d–f), challenging observation by Cai et al.39 Our data unambiguously show that TRIM72 is a negative feedback regulator of myogenesis by overexpression and knockdown of TRIM72 in skeletal muscle cells. For example, myogenesis was prevented in C2C12 and human primary skeletal muscle cells by adenoviral TRIM72 overexpression (Figure 2 and Supplementary Figure S3C–E). The C2C12 myogenesis was enhanced after the treatment of myotubes with four different si-TRIM72 oligomers (Figure 2 and Supplementary Figure S2). We also observed that myogenesis was highly enhanced in TRIM72−/− satellite cells compared to TRIM72+/+ satellite cells (Figure 5). In addition, TRIM72−/− mice have the increased cross-sectioned area of muscle fibers in tibialis anterior and soleus (unpublished data), indicating that TRIM72 is a real negative regulator of myogenesis.
Enhancement of skeletal muscle mass is known to be an excellent medical treatment for obesity and type II diabetes because insulin sensitivity and glucose uptake is highly increased in the enhanced skeletal muscles. Thus, the development of agents targeting TRIM72 could be a useful therapeutic approach to obesity and type II diabetes because TRIM72 inhibition might lead to increased insulin sensitivity in skeletal muscle by fortifying IGF-I-mediated IRS-1 activation.
Materials and Methods
Cell culture, animal, and antibodies
C2C12 cells were purchased from ATCC (Manassas, VA, USA) and grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 2% penicillin/streptomycin (WelGene, Daegu, Korea) and 10% fetal bovine serum (FBS; WelGene) in a 5% CO2 incubator at 37°C. Confluent C2C12 myoblasts were differentiated into myotubes by incubation with DMEM supplemented with 2% horse serum. Every 48 h, the myotubes were fed with fresh DMEM containing 2% horse serum.
Purified recombinant mouse TRIM72 peptides with amino-acid sequence 90–277 and 362–469 were used as antigens for rabbit polyclonal and mouse monoclonal antibodies, respectively. Anti-TRIM72 antibodies were generated by AbFrontier (Seoul, Korea). Various antibodies were from Upstate (Billerica, MA, USA) and BD Transduction Laboratories (San Jose, CA, USA) (anti-IRS-1, PI(3)K, p85, MyoD, myogenin, Akt, and phosphotyrosine), Santa Cruz Biotechnology (Santa Cruz, CA. USA) (IGF-IRβ, Myc, Flag, HA, Cav-3, Myf5, and pErk1/2), Sigma (St. Louis, MO, USA) (MyHC, Actin, and Flag), and Cell Signaling Technology (Danvers, MA, USA) (pAkt S473).
Satellite cell isolation
Hind limb muscles (10–12) were surgically removed from 4-week-old female mice or 10-week-old male mice, washed three times in Dulbecco's PBS, chopped in 0.025% of collagenase type I (Sigma) with fine scissors, incubated at 37°C for 90 min with stirring, and centrifuged at 300 × g for 15 min. The pellet was resuspended in 10 ml of complete medium (Ham's F10 supplemented with 20% v/v FBS) and seeded into 100-mm gelatin-coated dishes. Basic fibroblast growth factor was added twice a day at a concentration of 4 ng/ml. After 48 h, muscle debris was collected from medium after centrifugation at 300 × g for 5 min and resuspended in 15 ml of complete medium (F10/DMEM supplemented 20% FBS) containing 4 ng/ml of basic fibroblast growth factor. After preplating the suspended fragments for 20 min, they were transferred into new gelatin-coated dishes. On day 2, cultures were briefly treated with 0.025% collagenase at 37°C for the release of satellite cells. The satellite cells were centrifuged and resuspended in complete medium (F10/DMEM supplemented with 2% FBS and 5% Ultroser-G) containing 4 ng/ml of basic fibroblast growth factor. When cells were confluent, medium was changed with alpha minimum essential medium supplemented with 2% of horse serum for differentiation.
For the human primary skeletal muscle cell isolation, muscle samples were obtained from patients during surgical treatment for non-muscle problems of larynx and this study was approved by the institutional review board of Ajou University Hospital (AJIRB-CRO-05-087). The cells were isolated following a method of Foulstone et al.40 and maintained in F-10 medium with 20% FBS (Gibco-BRL, Carlsbad, CA, USA). Minimum Essential Medium supplemented with 2% horse serum was used for differentiation induction.
2-DE of detergent-resistant lipid rafts
Four 150 mm dishes of C2C12 myoblasts and myotubes were mixed with 1 ml of lysis buffer (25 mM HEPES (pH 6.5), 1% Triton X-100, 150 mM NaCl, 1 mM EDTA, 1 mM PMSF, and protease inhibitor cocktail), homogenized 20 times with a tight Dounce homogenizer (Kontes, Vineland, NJ, USA), and incubated for 30 min at 4°C. The extract was mixed with 1 ml of 80% sucrose, transferred to an SW41 centrifuge tube, and overlaid with 6 ml of 30% sucrose solution and 4 ml of 5% sucrose solution containing 25 mM HEPES (pH 6.5), 150 mM NaCl, and 1 mM EDTA. The discontinuous sucrose gradients were centrifuged for 18 h at 4°C at 39 000 r.p.m. The gradient was fractionated into 12 fractions from the bottom to the top. The lipid raft fractions were washed with washing buffer (25 mM HEPES (pH 6.5), 150 mM NaCl, and 1 mM EDTA) by ultracentrifugation (20 000 r.p.m., 30 min, 4°C), and suspended with 100 μl of resuspension buffer (9 M urea, 2 M thiourea, 4% CHAPS, protease inhibitor cocktail, and 1 mM EDTA). 2-DE, in-gel protein digestion, and LC-ESI-MS/MS were performed as previously described.12
Adenovirus preparation and infection
An adenoviral vector with TRIM72 was constructed using ViraPower Adenoviral Expression System (Invitrogen, Carlsbad, CA, USA) under the control of cytomegalovirus (CMV). Briefly, an adenoviral vector carrying the TRIM72 gene (Ad-TRIM72) was constructed using cDNA encoding the mouse TRIM72 under the control of CMV promoter. TRIM72 cDNA was cloned into Sal I/Not I-digested pENTR vector and then performed homologous recombination for introducing TRIM72 cDNA into the adenoviral destination vector as described in user manual. An adenoviral vector carrying the Escherichia coli LacZ reporter gene (Ad-EV) under the control of the CMV promoter was used as a control. Viral stocks were generated by infection of AD-293 cells and were concentrated by two rounds of cesium chloride ultracentrifugation. Cesium-banded virus stocks were dialyzed against 10 mM Tris (pH 7.9) and 1 mM MgCl2, aliquotted, and stored in −80°C until use. C2C12 myoblasts grown in six-well plates were infected by adenovirus with the dosage of 2 × 1010 viral particles per ml.
Plasmids, transient transfection, and luciferase assay
IRS-1 constructs (full, 1–3729; pleckstrin homology-phosphotyrosin binding domain, 1–900; middle region, 880–2608; C-terminal region, 2581–3729) and TRIM72 cDNA constructs (full, 1–1431; RING, 181–1431; CC, 367–1020; CC-SPRY, 367–1431; SPRY, 687–1431; ΔCC, TRIM72 mutant without 132–233) were generated by PCR and cloned into pCMV-Tag2B vector and pCMV-3Tag4A vector, respectively. For various deletion mutants of TRIM72 promoter, 1269, 1144, 883, and 556 bp upstream regions from ATG start codon were amplified by PCR. Forward primers were 5′-AAAGGTACCGAGCTTTGCAATATCTGG-3′ for 1269 bp fragment, 5′-GCCGGTACCGGAATAAATTGTAGGTCA-3′ for 1144 bp fragment, 5′-CCAGGTACCGAGTTAGCACCATTAGCG-3′ for 883 bp fragment, and 5′-TGCGGTACCGCCCTGAATCCCTGCCCT-3′ for 556 bp fragment and common reverse primer, 5′-CAGCAAGCTTGGTGAGCCTGGGAAGAGG-3′, was used for all the constructs. Site-directed mutagenesis of E1 and/or E2 boxes in 883 bp fragment was performed by PCR using the following primers: E1 mut forward, 5′-GCATCCACCCACTGC AATCTTGCCCTGAATCCC-3′; E1 mut reverse, 5′-GGGATTCAGGGCAAGATTGCAGTGGGTGGATGC-3′; E2 mut forward, 5′-CCACTATATGCCTATAACCTTTTGCATCCACCCACT-3′; and E2 mut reverse, 5′-AGTGGGTGGATGCAAAAGGTTATAGGCATATAGTGG-3′. These constructs were inserted into pGL3 basic vector (Promega, Madison, WI, USA). The open-reading frames of mouse MyoD, Myf5, myogenin, and Mrf4 were amplified by PCR and cloned into pcDNA3 vector (Invitrogen). Gene transfection was performed by using Geneporter (Genlantis, San Diego, CA, USA), Lipofectamin 2000 (Invitrogen), or electroporation (Digital Bio, Seoul, Korea) according to manufacturer's protocol. Luciferase activity was measured by using the luciferase assay system (Promega) in a Luminoskan Ascent (Thermo Labsystem, Waltham, MA, USA). We normalized the luciferase activity to the activity of coexpressed β-galactosidase. To calculate relative luciferase activity, we divided each normalized luciferase activity by that of empty vector-transfected cells.
RNA interference
siRNA oligomers targeting TRIM72 (si-TRIM72) were designed from Dharmacon (Lafayette, CO, USA), and scrambled oligomer (si-control) was obtained from Ambion (Austin, TX, USA). Target sequence of TRIM72 was 5′-AAGCACGCCUCAAGACACAGC-3′. For TRIM72 promoter study, scrambled (si-control), si-MyoD, and si-Akt oligomers were purchased from Santa Cruz Biotechnology. C2C12 myoblasts were transfected with si-control, si-TRIM72, si-MyoD, or si-Akt by using electroporation (Digital Bio) according to manufacturer's protocol.
Measurement of myogenic index
Myogenic index was determined as the ratio of the nuclei number in the cells containing two or more nuclei to the total nuclei number in hematoxylin-stained C2C12 myotubes or MyHC-stained satellite myotubes. C2C12 myotubes were washed in PBS, fixed with 10% formaldehyde in PBS for 10 min, and permeabilized with 0.5% Triton–X-100 in PBS for 5 min. These cells were then stained with 10% hematoxylin solution for 5 min and then washed in distilled water. The number of nuclei was measured at least from the six different fields observed under microscope (Olympus, Tokyo, Japan). Myogenesis of satellite cells was determined by MyHC immunofluorescence and DAPI staining. Fluorescence signal was observed under fluorescence microscope (Carl Zeiss, Medac, Wedel, Germany).
Western blotting, immunofluorescence, and immunoprecipitation
Western blotting and immunofluorescence were performed as previously described.12 For immunoprecipitation, cells were lysed in a buffer containing 20 mM Tris-HCl (pH 7.4), 137 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, 20 mM NaF, 10 mM Na4P2O7, 1 mM Na3VO4, 1% NP-40, 1 mM PMSF, and protease inhibitor cocktail. The whole-cell lysates (600 μg protein) were incubated with specific antibodies for 90 min and then with 50 μl of Protein A-sepharose or Protein G-agarose bead slurry for 90 min. The immunoprecipitates were analyzed by western blotting.
Northern blotting
Premade northern blot (MTN) containing mRNAs from different mouse organs was purchased from Clontech. After prehybridization, the premade northern blot was incubated with [α32P]-labeled TRIM72 probe (789–1419 bp fragment) for 2 h at 68°C in hybridization solution obtained from a kit (Clontech, Mountain View, CA, USA). The hybridized blot was washed, dried, and exposed to image plate. The signal was imaged by a BAS Reader (BAS-25000; Fuji Photo Film, Tokyo, Japan).
RT-PCR
Total RNA (1–2 μg) was isolated from C2C12 myotubes using Trizol reagent (Ambion), and were used as a template to prepare cDNA using M-MLV reverse transcriptase (Invitrogen). Semiquantitative PCR were performed using Taq polymerase (Genemed, South San Francisco, CA, USA) and the following primers. TRIM72 primers, 5′-TCCCTGTTGTCAGGCATCTAC-3′, and 5′-TTCTTCCACACCTGGAATTTG-3′; myogenin primers, 5′-GCTCAGCTCCCTCAACCAG-3′ and 5′-ATGTGAATGGGGAGTGGGA-3′; MyoD primers, 5′-CTCCTTTGAGACAGCAGACGACTT-3′ and 5′-AAATCGCATTGGGGTTTGAGCCTG-3′; Cav-3 primers, 5′-AGGACATTCACTGCAAGGAGA-3′ and 5′-CAGAAGGTGCGGATACACAGT-3′; MyHC primers, 5′-AGAAGGAGGAGGCAACTTCTG-3′ and 5′-ACATACTCATTGCCGACCTTG-3′; GAPDH primers, 5′-CTGCACCACCAACTGCTTAGC-3′ and 5′-CTTCACCACCTTCTTGATGTC-3′; actin primers, 5′-GAAGAGCTATGAGCTGCCTGA-3′ and 5′-CTCATCGTACTCCTGCTTGCT-3′.
Chromatin immunoprecipitation
C2C12 myoblasts and myotubes were fixed with 1% formaldehyde and lysed with a buffer containing 0.5% NP-40, 5 mM PIPES (pH 8.0), and 85 mM KCl. After brief microcentrifugation for 5 min at 5000 r.p.m., the pellet was further incubated with a nuclear lysis buffer containing 50 mM Tris-HCl (pH 8.0), 1% SDS, and 10 mM EDTA, sonicated and centrifuged for 10 min at 12 000 × g. The supernatant was used in ChIP assay by using anti-MyoD antibody as described.16 The immunoprecipitates were amplified by PCR with primers. The forward and reverse primers were 5′-AGGGAGTGGGTAGGACAGCTAAATAT-3′ and 5′-CAGGCTCAATGCAAGGGCAGGGA-3′, respectively.
Measurement of PI(3)K activity
The immunoprecipitates with an anti-p85 antibody were washed twice with buffer A (20 mM Tris-HCl (pH 7.4), 137 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, 20 mM NaF, 10 mM Na4P2O7, 1% NP-40, 3 mM benzamidine, 1 mM Na3VO4, and protease inhibitor cocktail) and reacted with 50 μl PI(3)K reaction buffer (10 mM Tris-HCl (pH 7.4), 100 mM NaCl, 1 mM EDTA, and 1 mM sodium vanadate) containing 60 μg of phosphoinositide (Sigma) and 20 μCi [γ-P32]. The phosphorylated lipids were separated by thin layer chromatography and imaged by a BAS Reader (BAS-25000; Fuji Photo Film).
Glucose uptake
C2C12 myotubes were incubated with 2 ml of Kreb's-Ringer phosphate buffer (128 mM NaCl, 1.4 mM CaCl2, 1.4 mM MgSO4, 5.2 mM KCl, and 10 mM Na2HPO4, pH 7.4) containing IGF-1 (200 ng/ml), 3H-2-deoxyglucose (10 μCi/ml), and glucose (5 mM). After incubation for 30 min at 37°C, the cells were washed three times with ice-cold PBS and solubilized with 0.3 N NaOH. The cell-associated radioactivity was determined by scintillation counting.
Generation and genotyping of TRIM72-disrupted mice
TRIM72-disrupted mice were generated by Macrogen (Seoul, Korea) using a standard procedure. Briefly, exons 1, 2, and 3 of the TRIM72 gene were replaced with a PGK-neo sequence within the targeting vector by homologous recombination. For screening of recombination-positive embryonic stem cells, genomic DNA was digested with BamHI and analyzed by Southern blotting. Genotyping was performed by PCR, using specific primers for wild-type and knockout alleles. The primers are as follows: 5′-CCAGCCCATGACTGAATTTT-3′/5′-GCACCGCTACAGTCTTCTCC-3′ for the wild-type allele, and 5′-TGATATTGCTGAAGAGCTTGG-3′/5′-ACAAAGGTGGCCTGAGTGATC-3′ for the knockout allele. Mice were housed in plastic cages on a 12:12 h light/dark photoperiod with free access to water and food. Animals were handled according to the Principles of Laboratory Animal Care (NIH Publication No. 85-23, revised 1985).
Abbreviations
- MURF-1:
-
muscle-specific RING finger-1
- 2-DE:
-
two-dimensional electrophoresis
- Cav-3:
-
caveolin-3
- FOXO:
-
forkhead box O
- GSK3β:
-
glycogen synthase kinase 3β
- IGF:
-
insulin-like growth factor
- IGFR:
-
insulin-like growth factor receptor
- IRS-1:
-
insulin receptor substrate-1
- LC-ESI-MS/MS:
-
liquid chromatography-electrospray ionization-tandem mass spectrometry
- MyHC:
-
myosin heavy chain
- mTOR:
-
mammalian target of rapamycin
- PI(3)K:
-
phosphoinositide 3-kinase
- RING:
-
really interesting new gene
- RT-PCR:
-
reverse transcription-polymerase chain reaction
- SPRY:
-
SPla and RYanodine receptor
- TRIM72:
-
tripartite motif 72
References
Simons K, Toomre D (2000) Lipid rafts and signal transduction. Nat Rev Mol Cell Biol 1: 31–39.
Jacobson K, Mouritsen OG, Anderson RG (2007) Lipid rafts: at a crossroad between cell biology and physics. Nat Cell Biol 9: 7–14.
Brown DA, Rose JK (1992) Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell 68: 533–544.
Parton RG (2003) Caveolae – from ultrastructure to molecular mechanisms. Nat Rev Mol Cell Biol 4: 162–167.
Hill MM, Bastiani M, Luetterforst R, Kirkham M, Kirkham A, Nixon SJ et al. (2008) PTRF-Cavin, a conserved cytoplasmic protein required for caveola formation and function. Cell 132: 113–124.
Minetti C, Sotgia F, Bruno C, Scartezzini P, Broda P, Bado M et al. (1998) Mutations in the caveolin-3 gene cause autosomal dominant limb-girdle muscular dystrophy. Nat Genet 18: 365–368.
Viola A, Gupta N (2007) Tether and trap: regulation of membrane-raft dynamics by actin-binding proteins. Nat Rev Immunol 7: 889–896.
Kim K-B, Kim B-W, Choo H-J, Kwon Y-C, Ahn B-Y, Choi J-S et al. (2009) Proteome analysis of adipocyte lipid rafts reveals that gC1qR plays essential roles in adipogenesis and insulin signal transduction. Proteomics 9: 2373–2382.
Foster LJ, De Hoog CL, Mann M (2003) Unbiased quantitative proteomics of lipid rafts reveals high specificity for signaling factors. Proc Natl Acad Sci USA 100: 5813–5818.
Kim KB, Kim SI, Choo HJ, Kim JH, Ko YG (2004) Two-dimensional electrophoretic analysis reveals that lipid rafts are intact at physiological temperature. Proteomics 4: 3527–3535.
Bae TJ, Kim MS, Kim JW, Kim BW, Choo HJ, Lee JW et al. (2004) Lipid raft proteome reveals ATP synthase complex in the cell surface. Proteomics 4: 3536–3548.
Kim KB, Lee JW, Lee CS, Kim BW, Choo HJ, Jung SY et al. (2006) Oxidation-reduction respiratory chains and ATP synthase complex are localized in detergent-resistant lipid rafts. Proteomics 6: 2444–2453.
Glass DJ (2003) Molecular mechanisms modulating muscle mass. Trends Mol Med 9: 344–350.
Glass DJ (2003) Signalling pathways that mediate skeletal muscle hypertrophy and atrophy. Nat Cell Biol 5: 87–90.
Erbay E, Park I-H, Nuzzi PD, Schoenherr CJ, Chen J (2003) IGF-II transcription in skeletal myogenesis is controlled by mTOR and nutrients. J Cell Biol 163: 931–936.
Wilson EM, Rotwein P (2006) Control of MyoD function during initiation of muscle differentiation by an autocrine signaling pathway activated by insulin-like growth factor-II. J Biol Chem 281: 29962–29971.
Musarò A, McCullagh K, Paul A, Houghton L, Dobrowolny G, Molinaro M et al. (2001) Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat Genet 27: 195–200.
Rommel C, Clarke BA, Zimmermann S, Nunez L, Rossman R, Reid K et al. (1999) Differentiation stage-specific inhibition of the Raf-MEK-ERK pathway by Akt. Science 286: 1738–1741.
Murgia M, Serrano AL, Calabria E, Pallafacchina G, Lomo T, Schiaffino S (2000) Ras is involved in nerve-activity-dependent regulation of muscle genes. Nat Cell Biol 2: 142–147.
Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R et al. (2001) Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol 3: 1014–1019.
Rommel C, Bodine SC, Clarke BA, Rossman R, Nunez L, Stitt TN et al. (2001) Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat Cell Biol 3: 1009–1013.
Hribal ML, Nakae J, Kitamura T, Shutter JR, Accili D (2003) Regulation of insulin-like growth factor-dependent myoblast differentiation by Foxo forkhead transcription factors. J Cell Biol 162: 535–541.
Sandri M, Sandri C, Gilbert A, Skurk C, Calabria E, Picard A et al. (2004) Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 117: 399–412.
Huo H, Guo X, Hong S, Jiang M, Liu X, Liao K (2003) Lipid rafts/caveolae are essential for insulin-like growth factor-1 receptor signaling during 3T3-L1 preadipocyte differentiation induction. J Biol Chem 278: 11561–11569.
Panetta D, Biedi C, Repetto S, Cordera R, Maggi D (2004) IGF-I regulates caveolin 1 and IRS1 interaction in caveolae. Biochem Biophys Res Commun 316: 240–243.
Arcaro A, Aubert M, Espinosa del Hierro ME, Khanzada UK, Angelidou S, Tetley TD et al. (2007) Critical role for lipid raft-associated Src kinases in activation of PI3K-Akt signalling. Cell Signal 19: 1081–1092.
Sui Z, Kovacs AD, Maggirwar SB (2006) Recruitment of active glycogen synthase kinase-3 into neuronal lipid rafts. Biochem Biophys Res Commun 345: 1643–1648.
Ozato K, Shin DM, Chang TH, Morse III HC (2008) TRIM family proteins and their emerging roles in innate immunity. Nat Rev Immunol 8: 849–860.
Witt SH, Granzier H, Witt CC, Labeit S (2005) MURF-1 and MURF-2 target a specific subset of myofibrillar proteins redundantly: towards understanding MURF-dependent muscle ubiquitination. J Mol Biol 350: 713–722.
McElhinny AS, Perry CN, Witt CC, Labeit S, Gregorio CC (2004) Muscle-specific RING finger-2 (MURF-2) is important for microtubule, intermediate filament and sarcomeric M-line maintenance in striated muscle development. J Cell Sci 117: 3175–3188.
Kudryashova E, Kudryashov D, Kramerova I, Spencer MJ (2005) Trim32 is a ubiquitin ligase mutated in limb girdle muscular dystrophy type 2H that binds to skeletal muscle myosin and ubiquitinates actin. J Mol Biol 354: 413–424.
Chen D, Gould C, Garza R, Gao T, Hampton RY, Newton AC (2007) Amplitude control of protein kinase C by RINCK, a novel E3 ubiquitin ligase. J Biol Chem 282: 33776–33787.
Cai C, Masumiya H, Weisleder N, Matsuda N, Nishi M, Hwang M et al. (2009) MG53 nucleates assembly of cell membrane repair machinery. Nat Cell Biol 11: 56–64.
Meroni G, Diez-Roux G (2005) TRIM/RBCC, a novel class of ‘single protein RING finger’ E3 ubiquitin ligases. Bioessays 27: 1147–1157.
Forcales SV, Puri PL (2005) Signaling to the chromatin during skeletal myogenesis: novel targets for pharmacological modulation of gene expression. Semin Cell Dev Biol 16: 596–611.
Lee CH, Inoki K, Guan KL (2007) mTOR pathway as a target in tissue hypertrophy. Annu Rev Pharmacol Toxicol 47: 443–467.
Wilson EM, Hsieh MM, Rotwein P (2003) Autocrine growth factor signaling by insulin-like growth factor-II mediates MyoD-stimulated myocyte maturation. J Biol Chem 278: 41109–41113.
Serra C, Palacios D, Mozzetta C, Forcales SV, Morantte I, Ripani M et al. (2007) Functional interdependence at the chromatin level between the MKK6/p38 and IGF1/PI3K/AKT pathways during muscle differentiation. Mol Cell 28: 200–213.
Cai C, Masumiya H, Weisleder N, Pan Z, Nishi M, Komazaki S et al. (2008) MG53 regulates membrane budding and exocytosis in muscle cells. J Biol Chem 284: 3314–3322.
Foulstone EJ, Savage PB, Crown AL, Holly JM, Stewart CE (2003) Role of insulin-like growth factor binding protein-3 (IGFBP-3) in the differentiation of primary human adult skeletal myoblasts. J Cell Physiol 195: 70–79.
Acknowledgements
We thank M-S Kim for CA-FOXO gene and E-J Choi for CA-GSK3β gene. We also thank Dr. Guy A Thompson for critical discussions. This work was supported by grants from the Center for New Drug Target Discovery of the Korea Ministry of Science and Technology (2006-02795), Korea Research Foundation (KRF-2005-C00358), and Korean Ministry of Public Health and Welfare (A090597) to Y-G Ko.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by G Cossu
Supplementary Information accompanies the paper on Cell Death and Differentiation website
Supplementary information
Rights and permissions
About this article
Cite this article
Lee, C., Yi, JS., Jung, SY. et al. TRIM72 negatively regulates myogenesis via targeting insulin receptor substrate-1. Cell Death Differ 17, 1254–1265 (2010). https://doi.org/10.1038/cdd.2010.1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2010.1
Keywords
This article is cited by
-
Structural basis for TRIM72 oligomerization during membrane damage repair
Nature Communications (2023)
-
The Pivotal Role of Mitsugumin 53 in Cardiovascular Diseases
Cardiovascular Toxicology (2021)
-
An MG53-IRS1-interaction disruptor ameliorates insulin resistance
Experimental & Molecular Medicine (2018)
-
Heart Failure and MEF2 Transcriptome Dynamics in Response to β-Blockers
Scientific Reports (2017)
-
Evaluation of common variants in MG53 and the risk of type 2 diabetes and insulin resistance in Han Chinese
SpringerPlus (2016)
