
Abstract
Catecholamines regulate the β -adrenoceptor/cyclic AMP-regulated protein kinase A (cAMP/PKA) pathway. Deregulation of this pathway can cause apoptotic cell death and is implicated in a range of human diseases, such as neuronal loss during aging, cardiomyopathy and septic shock. The molecular mechanism of this process is, however, only poorly understood. Here we demonstrate that the β -adrenoceptor/cAMP/PKA pathway triggers apoptosis through the transcriptional induction of the pro-apoptotic BH3-only Bcl-2 family member Bim in tissues such as the thymus and the heart. In these cell types, the catecholamine-mediated apoptosis is abrogated by loss of Bim . Induction of Bim is driven by the transcriptional co-activator CBP (CREB-binding protein) together with the proto-oncogene c-Myc. Association of CBP with c-Myc leads to altered histone acetylation and methylation pattern at the Bim promoter site. Our findings have implications for understanding pathophysiology associated with a deregulated neuroendocrine system and for developing novel therapeutic strategies for these diseases.
Similar content being viewed by others

Main
Cyclic adenosine monophosphate (cAMP) is a second messenger that is highly conserved throughout evolution. In metazoans, its primary role is to act as an intracellular carrier of metabolic information, regulating hormonal responses,1 in triggering apoptotic cell death and in regulating ontogeny.2, 3 Studies with transgenic mouse models have revealed that deregulation of the cyclic AMP-regulated protein kinase A (cAMP/PKA) pathway can cause apoptosis.4 Specifically, β-adrenergic receptor (βAR) activation by catecholamines or autoantibodies, as seen in Grave’s disease,5 can cause PKA-mediated cardiomyocyte apoptosis, resulting in heart failure (HF).6 Furthermore, administration of catecholamines, such as epinephrine, is often the last resort for treatment of patients suffering from septic shock,7 and this is associated with profound alterations in immune function, similar to those observed in haemorrhagic shock where all lymphocyte subsets are decreased owing to excessive apoptosis.8 Similarly, loss of dopamine receptor containing neurons during aging has been attributed to the apoptosis-inducing effects of the catecholamine, dopamine9 and chronic stress-induced immune modulation has been attributed to increased levels of circulating epinephrine.10
The molecular mechanisms of cAMP/PKA pathway-mediated apoptosis are only poorly defined. The BH3-only Bcl-2 family protein Bim is considered to be an essential initiator of apoptosis in a wide variety of physiological settings, including deregulated calcium flux, growth factor withdrawal,11 endoplasmic reticulum stress12 and T cell receptor13 as well as B cell receptor activation14 in autoreactive lymphocytes. Downregulation of Bim appears to be a common denominator in many cell survival signalling pathways in cancers, and many anticancer therapeutics kill cancer cells by inducing Bim expression.15, 16 We have previously reported that the cAMP/PKA pathway can regulate Bim protein levels by phosphorylation and stabilization.17 However, the present work demonstrates that in many cell types and tissues, transcriptional induction of Bim by the cAMP/PKA pathway has a major role in cell death regulation.
Here we elucidate the molecular mechanism of the cAMP/PKA-triggered apoptotic pathway downstream of βAR activation. We demonstrate that PKA activation results in the transcriptional induction of the pro-apoptotic BH3-only Bcl-2 family gene Bim and subsequent Bim-dependent apoptosis in diverse cell types, including thymocytes and cardiomyocytes. We also provide proof for the engagement of the Bim promoter by the proto-oncogene c-Myc, together with the transcriptional co-activator CREB (cAMP response element-binding protein)-binding protein (CBP). Our results shed light on the role of increased sympathetic nerve activity and apoptosis in the myocardium that can lead to HF18, in the development of the thymus during embryogenesis19 and in stress-induced thymic atrophy and immune modulation.20
Results
cAMP/PKA activation induces BIM in a variety of tissues
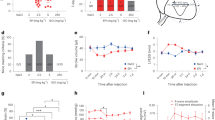
A critical role for the pro-apoptotic BH3-only Bcl-2 family member Bim (Bcl2L11) in cAMP/PKA-triggered apoptosis has been reported in the S-49 T-cell lymphoma line.21 To test the generality of this phenomenon, we used a variety of systems to induce PKA activity. PKA exists as a tetrameric holoenzyme composed of two catalytic (C) and two regulatory (R) subunits. Binding of cAMP causes the dissociation of the holoenzyme into its constituent subunits, that is, PKA regulatory subunit 1a (Prkar1a) and PKACα However, cAMP regulation of PKA can be bypassed by ectopic overexpression of PKACα.22 4-hydroxytamoxifen (4-OHT)-induced expression of PKACα HA-tagged) in mouse embryonic fibroblasts (MEFs) resulted in the induction of Bim, both at the protein and mRNA levels (Figure 1a). Similarly, βAR stimulation by agonists, such as isoproterenol, leads to cAMP flux and PKA activation.21 Consistent with this, treatment of MEFs with isoproterenol led to a robust induction of Bim both at the protein and mRNA levels (Figure 1b and Supplementary Figure S1d).

cAMP/PKA activation induces Bim in a variety of tissues. (a) MEFs expressing PKACα under 4-OHT regulation. First two lanes are samples from a clone, which expresses very little PKACα used as control. pCREB is the surrogate marker for PKA activation. (b) MEFs treated with isoproterenol for various time points and analyzed by western blot or by qPCR. Nur77 transcripts are used as a marker for PKA activation in the qPCR analysis. (c and d) Thymocytes in FTOC or cardiomyocytes in culture were treated with isoproterenol and rolipram (10 μM each) and analyzed for various transcripts by qPCR. (e) β2AR transgenic mouse (β2AR expressed under MHC promoter or MHCα) and littermate control on C57BL/6 background (at 6 months) were analyzed by echocardiogram, TUNEL staining for apoptosis or by Masson’s trichrome staining for collagen accumulation. (f) LV sections were stained with Bim-specific antibodies or the antibodies pre-incubated with the peptide epitope against which the antibodies were raised to show the specificity of the staining. (g) RT-qPCR analysis of the heart tissues for Bim transcripts and (h) western blot analysis of the transgenic heart tissues with myocardial infarct (MI) as a positive control. Error bars: ±S.E.M., n=3, one-tailed T-tests. #P=0.004; *P=0.023 versus untreated control (b); #P=0.002–0.0012 versus untreated control (c); #P=0.0014 versus untreated control (d); #P=0.013 versus wild-type (WT) control
The primary and secondary lymphoid organs, such as the developing thymi as well as the spleen and lymph nodes, are innervated by the autonomic, mainly sympathetic, nervous system, allowing neuroimmune modulation.23 Postnatal infection and stress can cause profound involution of the thymus gland, and this is attributed to increased secretion of catecholamines by the sympathetic nervous system.24 As Bim is critical for the killing of thymocytes by diverse apoptotic stimuli,11 we tested whether thymocytes could upregulate Bim in response to catecholamines. Treatment of thymocytes in foetal thymic organ culture (FTOC) with isoproterenol resulted in a robust induction of Nur77, a marker for PKA activation25 and Bim, whereas the expression of other BH3-only genes, such as Puma (Bbc3) and Noxa remained unchanged (Figure 1c).
β-adrenergic stimulation leads to PKA-mediated apoptosis in adult cardiomyocytes.26 Accordingly, isoproterenol treatment induced Nur77 and Bim expression in cardiomyocytes isolated from adult mice by Langendorff perfusion (Figure 1d). Similar results were obtained in in vivo mouse models. Similar to what was seen in β1AR transgenic animals,27 cardiomyocyte-specific transgenic overexpression of β2AR led to cardiomyocyte apoptosis, hypertrophy and ultimately HF.28 These transgenic mice had enlarged hearts, dilated left ventricles (LV) and increased fibrosis, as revealed by echocardiogram, TUNEL and collagen staining (Figure 1e and Supplementary Figure S1a). Immunohistochemical staining of heart sections demonstrated that hearts in these transgenic mice expressed higher levels of Bim compared with hearts from wild-type (WT) mice (Figure 1f). Quantitative PCR (qPCR) and western blot analyses of these heart tissues indicated that Bim was induced in these mice both at the mRNA and protein levels (Figures 1g and h). The heart is richly innervated by sympathetic nerves, which regulate cardiac function by the release of catecholamines that act on βARs. Excessive catecholamine levels are correlated with failing myocardium.29 Consistent with this notion, induction of myocardial infarction by left coronary artery ligation resulted in increased levels of both adrenaline and nor-adrenaline in the serum of C57BL/6 mice, and this was accompanied by increased Bim protein expression (Supplementary Figures S1b and c). Similar to what we observed in thymocytes, this induction was specific to Bim, as we did not detect any induction of other BH3-only proteins, such as Puma (Supplementary Figure S1c).
Loss of Bim inhibits cAMP/PKA-induced apoptosis
Consistent with the reports from T hybridoma cells,21 our results show that Bim is a downstream target of the cAMP/PKA pathway in diverse tissues. We therefore examined whether Bim was required for PKA-induced apoptosis. Apart from the sympathetic nervous system, immune cells themselves can synthesize and metabolize catecholamines and stimulate βARs. Immune cells are therefore considered to be ‘a new, diffusely distributed adrenergic organ’.4 Catecholamines regulate lymphocyte proliferation, differentiation and apoptosis via an autocrine loop.30 Therefore, we tested whether loss of Bim could protect thymocytes from βAR-mediated apoptosis in FTOC. Treatment of FTOC with isoproterenol resulted in Bim protein induction (Supplementary Figure S2a) accompanied by apoptosis of the WT thymocytes. Loss of Bim, not loss of Puma rendered thymocytes completely resistant to this treatment (Figures 2a and b). This was not because of attenuated PKA activation in Bim −/− cells, as treatment with isoproterenol activated PKA to similar extent in both Bim −/− and WT cells, as indicated by the increase in phospho-CREB (pCREB) and cAMP levels (Supplementary Figure S2a, lanes 2 and 4 and Figure 2c). (Constitutive levels of both pCREB and cAMP levels were higher in Bim −/− cells; this might be because of the survival of thymocytes with high cAMP/PKA activity in the absence of Bim.)

cAMP/PKA-mediated apoptosis is Bim dependent. (a and b) Analysis of thymocytes apoptosis in FTOC in response to βAR activation. (c) cAMP measurement in thymocytes from WT and Bim−/− mice in response to βAR activation. (d) Representative echocardiographic 2D short-axis view of the LV of mice injected with isoproterenol (15 mg/kg per day i.p. for 8 days) compared with untreated controls (UT). (e and f) Measurement of fractional shortening and fractional area change in the isoproterenol-treated mice compared with control. Error bars: ±S.E.M., n=3–5. #P=0.0053–0.0078 (b and c); n=5–7, #P=0.0067–0.0094 (e and f); *P=0.432–0.562; one-tailed T-tests
In an acute mouse model of HF, injection of isoproterenol (15 mg/kg per day for 7 days) causes ventricular remodelling, leading to cardiac hypertrophy.31 Although isoproterenol injection resulted in impaired contractile function of the LV (significantly lower fractional shortening and lower fractional area change) in the WT mice, the Bim−/− mice did not develop this pathology (Figures 2d–f). Accordingly, many more TUNEL+ (apoptotic) cells were seen in WT mice compared with the Bim−/− animals (Supplementary Figure S2b). Similarly, whereas cardiomyocytes isolated from adult WT mice by Langendorff perfusion underwent substantial apoptosis after treatment with isoproterenol, those from Bim−/− mice were resistant (Supplementary Figure S2c).
Apart from rescuing the catecholamine-induced pathology, Bim ablation also resulted in constitutive physiological hypertrophy of the heart. This is characterized by increased weight of LV and whole heart, and this could be attributed to increased size of cardiomyocytes (Figures 3a and b and Supplementary Figure S2d). Furthermore, gene expression analysis of WT and Bim−/− heart tissues indicated that the cardiac hypertrophy is not associated with any pathology, as we did not observe any of the pathological hypertrophy-associated gene induction in the Bim−/− hearts (Figure 3c).

Bim ablation leads to increased cardiomyocyte size. (a) Wheat germ agglutinin staining of heart section from mice treated with isoproterenol (ISO). (b and d) Cardiomyocyte size determination from the WT and Bim−/− heart sections. (c) qPCR analysis of pathology-associated gene expression in the WT and Bim−/− hearts. Error bars: ±S.E.M., n=5–7, #P=0.0009; *P=0.2309–0.8809; paired one-tailed T-tests
Prkar1a deletion induces apoptosis and this can be inhibited by loss of Bim
The discovery that the exchange protein activated by cAMP (Epac) is a direct target of cAMP with an affinity comparable to the regulatory subunit of PKA32 questions the long-held notion that PKA is the only direct downstream target of cAMP. To rule out that Epac is critical for the induction of Bim, we used a genetic model in which PKA can be activated without inducing cAMP flux or adenylyl cyclase activation. Deletion of the ubiquitously expressed regulatory subunit of PKA, R1α (Prkar1a), leads to constitutive activation of PKA.33 Accordingly, MEFs from Prkar1afl/fl mice were infected with lentiviruses expressing 4-OHT-inducible CRE.34 This resulted in acute loss of R1α with consequent PKA activation (as seen by increased levels of phospho-PKA substrates) and in a robust induction of Bim both at the mRNA and protein levels (Figure 4a). Induction of Bim triggered by R1α deletion resulted in a substantial reduction in the clonogenic survival of these MEFs. Remarkably, this could be prevented by the loss of Bim (Figure 4b). This was also seen in short-term survival assay as well as when Prkar1a-deficient MEFs were treated with isoproterenol (Figure 4c).

Activation of cAMP/PKA by targeted deletion of Prkar1a in MEFs induces Bim-mediated apoptosis. (a) CRE-mediated deletion of Prkar1a in MEFs using 4-OHT-inducible lentiviral system induces Bim both at protein and RNA levels. (b) 100 cells of various genotypes (Prkar1a fl/fl; Prkar1a −/−; Prkar1a fl/fl, Bim−/− and Prkar1a−/−, Bim−/−) were plated for clonogenic survival on six-well plates and stained with crystal violet and (c) MEFs of various genotypes as shown were treated with 4-OHT to induce CRE expression, and then treated with isoproterenol (10 μM) in serum-free medium and apoptosis was measured. Error bars: ±S.E.M., n=3, #P=0.006–0.042 (b); # P=0.072 (c); one-tailed T-tests
c-Myc regulates BIM transcription during βAR stimulation
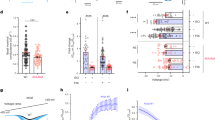
To identify the transcription factor(s) involved in βAR stimulation-mediated induction of Bim, we used the MEFs with 4-OHT-inducible PKACα expression (Figures 1a and 5a, inset). Transient transfection of these cells with a Bim promoter-luciferase reporter resulted in a robust induction of luciferase upon addition of 4-OHT. Using this assay, we mapped the Bim promoter and identified a conserved hexa-nucleotide region 140 bp upstream of the transcription start site, which was essential for PKA-mediated Bim upregulation (Figure 5a). This site potentially represents a non-canonical E-box capable of binding the proto-oncogene c-Myc,35 and binding of Myc to this site could indeed be confirmed in electrophoretic mobility shift assays (EMSA) (Supplementary Figure S3e). A critical role for c-Myc in Bim mRNA upregulation was first reported in the Eμ-Myc mouse pre-B/B lymphoma model. Bim mRNA and Bim protein levels were elevated in the apoptosis-prone B lymphoid cells of Eμ-MYC transgenic mice, and Bim-deficient Eμ-Myc mice had increased numbers of sIgM+ B cells.36 However, a direct relationship between c-Myc and Bim transcription is yet to be established. Expression of c-Myc under 4-OHT-regulation in MEFs resulted in a robust induction of Bim at both the mRNA and protein levels as, for example, seen in the luciferase assay (Figure 5b). This transcriptional induction of the Bim reporter construct could be abrogated either by mutating the conserved E-Box or by using the PKA inhibitor H-89 (Figure 5b). Furthermore, βAR stimulation-triggered induction of the Bim reporter in MEFs could also be abrogated by 10058-F4,37 a specific inhibitor of c-Myc–Max interaction. This indicates that Myc–Max heterodimerization is crucial for this transcriptional upregulation of Bim (Figure 5c). Consistent with these results, acute loss of c-Myc by CRE-mediated deletion in c-Mycfl/fl MEFs resulted in failure to induce Bim in response to βAR activation, although both c-Mycfl/fl and c-Myc−/− MEFs were equally capable of activating the βAR pathway, as evidenced by the increase in p-CREB levels (Figure 5d).

c-Myc regulates BIM transcription during cAMP/PKA activation. (a) Mapping of the Bim promoter by luciferase assay using MEFs expressing PKACα under 4-OHT-regulation. Western blot showing Bim induction is shown in the inset. The sequence identified is shown in blue (WT) or red (mutant). (b) Confirmation of c-Myc regulation in reporter assays using MEFs expressing c-Myc under 4-OHT-regulation as in (a). H-89 is used as the PKA inhibitor. (c) Myc-Max inhibitor, 10058-F4 blocks Bim induction in luciferase assays and in western blots (inset). (d) Deletion of c-MYC in MEFs abrogates βAR-mediated Bim induction. pCREB acts as the marker for cAMP/PKA activation. Deletion of c-Mycfl/fl allele is shown by PCR. (e) Whole genome ChIP analysis of c-Myc binding to the Bim promoter in different human cell lines. Base overlap is shown on the X axis. The conserved c-Myc-binding sequence on the Bim promoter is shown at the bottom. (f) MEFs from WT and the Bim promoter knock-in mouse where c-Myc-binding sites were mutated, were infected with lentiviruses expressing c-Myc under 4-OHT-regulation (top panel) or were treated with isoproterenol (bottom panel). The knock-in cells show lack of Bim induction. Non-specific band (indicated by asterisk) acts as the loading control. Error bars: ±S.E.M., n=3, #P=0.007–0.016 versus untreated controls (a–c); one-tailed T-tests
Whole genome chromatin immunoprecipitation (ChIP) and Next Generation Sequence analysis data generated as part of the ENCODE consortium by the Iyer lab (University of Texas, Austin) demonstrated that the Bim promoter was occupied by c-Myc in a variety of human cell lines (Figure 4e; http://www.genome.ucsc.edu/cgi-bin/hgTracks?hgS_doOtherUser=submit&hgS_otherUserName=Vishy&hgS_otherUserSessionName=bim_Myc). The c-Myc-binding region identified by ChIP spanned the sequence identified in the luciferase reporter assays corroborating our results (Figure 5e). To validate and expand these findings, we generated a knock-in mouse strain in which the conserved c-Myc-binding sites within the Bim promoter were mutated (Supplementary Figure S3a). In MEFs derived from these mutant mice, Bim transcription was not upregulated upon enforced c-Myc expression or βAR activation (Figure 5f and Supplementary Figure S3b). This demonstrates that c-Myc is essential for βAR-mediated Bim induction.
CBP is a required cofactor for c-Myc in Bim transcriptional induction
The data so far suggest that the transcription factor c-Myc is the critical driver of Bim transcriptional induction in response to cAMP/PKA stimulation. However, the link between PKA and c-Myc remains to be established. The only published report linking c-Myc directly with PKA is the transcriptional induction of the PKA catalytic subunit beta.38 We first examined whether c-Myc was a direct phosphorylation substrate of PKA, as c-Myc protein has canonical PKA phosphorylation sites (158RKDS161 and 276KRSES280 as predicted by the IMP Bioinformatics Group algorithm, Austria). Indeed, both WT and S279A mutant c-Myc could be readily phosphorylated by PKA in vitro, whereas S159A mutant c-Myc could not be phosphorylated (Supplementary Figure S3c). This mutation did, however, not have any discernable effect on the DNA-binding activity of c-Myc, as shown by EMSA and transcriptional activation of the Bim reporter construct or on the induction of Bim in MEFs when stably expressed, using 4-OHT-inducible lentiviral system (Supplementary Figures S3d–f). The transcriptional activity of c-Myc can be augmented by the recruitment of the cofactor CBP.39 We therefore tested whether such a process is critical for PKA-induced c-Myc transcriptional activity by RNAi-mediated knockdown of Cbp in MEFs. Interestingly, this resulted in the blockage of Bim induction, although both the WT and Cbp-knockdown cells were equally responsive to the βAR agonist, as reflected by CREB phosphorylation (Figure 6a). Furthermore, induction of the Bim luciferase reporter was markedly impaired in Cbp-knockdown cells (Figure 6b). Consequently, these cells were resistant to βAR agonist-induced apoptosis (Supplementary Figure S4a). To prove that CBP regulates Bim expression through c-Myc, we used the Bim-luciferase reporter assay. Transfection of MEFs with CBP and the WT Bim promoter-luciferase construct resulted in a robust induction of this reporter in response to treatment with isoproterenol, whereas such induction was not seen with the reporter in which the c-Myc-binding site had been mutated (Figure 6c).

CBP is the co-factor for c-Myc for Bim induction. (a and b) RNAi knockdown of CBP in MEFs results in lack of Bim induction up on βAR activation in western blot and in luciferase assays. (c) CBP-mediated Bim induction is dependent on intact c-Myc-binding sites on the Bim promoter. (d) HAT-deficient mutants of CBP fail to induce Bim. Acetylation of c-Myc by CBP or by the mutant variants in 293T cells is shown in the inset. (e) LK-2 cells that lack functional CBP respond to βAR signalling by inducing Bim protein in CBP-dependent manner. Error bars: ±S.E.M., n=3, #P=0.0022 versus untreated control (b); #P=0.008 versus isoproterenol-treated CBP control (c); #P=0.064 versus CBP control (e); paired one-tailed T-tests
Inactivating mutations in CBP are frequently found in acute lymphoblastic leukemia and in diffuse large cell B cell lymphomas. A large proportion of these mutations reside in the histone acetyltransferase (HAT) domain of CBP.40 Moreover, epigenetic silencing of Bim is a feature in some types of B cell lymphoma.41 We therefore examined whether HAT-deficient mutants of CBP were impaired in their ability to transcriptionally induce Bim. WT CBP could readily acetylate c-Myc in 293T cells but the HAT-deficient mutants H1451C or Y1234C40 had lost this ability (Figure 6d). Consistent with this, WT CBP could induce the Bim reporter, whereas the mutants failed to do so (Figure 6d). These results were further corroborated in the human lung cancer-derived cell line LK-2, which does not have a functional CBP protein owing to a deletion of exon 3 of the Cbp gene.42 βAR activation could readily induce Bim expression in the osteosarcoma cell line (143B) or B cell lymphoma cell line (Ramos), both of which have functional CBP, but not in CBP-deficient LK-2 cells (Supplementary Figure S4b). However, introduction of WT Cbp could restore βAR stimulation-triggered Bim induction in these cells (Figure 6e and Supplementary Figures S4c and d). These results demonstrate an essential role for CBP in c-Myc-mediated Bim induction during βAR activation.
Epigenetic regulation of Bim during βAR signalling
The Bim gene is subject to epigenetic changes both by histone modification and by DNA methylation. In many cancers, such as multiple myeloma, treatment with histone deacetylase inhibitors increases Bim expression and triggers apoptosis in a manner that is dependent (at least in part) on Bim.43 In chronic myeloid leukemia, changes in Bim promoter methylation status are associated with differences in Bim gene expression.44 As CBP is a cofactor in c-Myc-regulated Bim expression, and CBP mutations in the HAT domain are associated with tumor development,40 we investigated whether epigenetic changes in the Bim promoter occur during βAR signalling. Bisulphite sequence analysis of the Bim promoter from βAR responsive Ramos cells and the non-responsive LK-2 (CBP negative) cells showed a significant increase in CpG island methylation in the non-responsive cells. This could be partially reversed by the ectopic expression of CBP from a lentiviral vector (Figure 7a). Finally, ChIP analysis using MEFs expressing the catalytic subunit of PKA under 4-OHT regulation showed that both c-Myc and CBP bind constitutively to the Bim promoter, irrespective of the PKA activation status (Figure 7b). This is consistent with the whole genome ChIP analysis (Figure 5e), which showed that c-Myc could constitutively bind to the Bim promoter in different cell lines. However, histone 4 acetylation at the Bim promoter occurred specifically during PKA activation, corroborating the role of CBP in histone modification (Figure 7b last lane). These results establish a role for CBP in the cAMP/PKA pathway in inducing epigenetic changes in the Bim gene.

Bim promoter region is subject to epigenetic changes during βAR signalling. (a) Bisulphite sequencing analysis of RAMOS, LK-2 and LK-2 cells expressing full-length CBP using primers flanking the region between −1513 and −948 (from the transcription initiation site). The numbers shown are the potential CpG methylation sites. (b) ChIP analysis of the Bim promoter in MEFs expressing PKACα using antibodies as indicated. (c) A model for Bim induction by cAMP/PKA pathway. PKA activation of CBP bound to the Bim promoter through its interaction with c-Myc results in histone 4 acetylation, leading to Bim transcriptional induction
Discussion
Increased sympathetic nervous activity in the myocardium is a classical feature of patients with HF.18 Similarly, catecholamines, the primary transmitter of the sympathetic nervous system, regulate lymphocyte homeostasis by both autocrine and paracrine signalling.4 In cardiac βAR signalling, it is well established that chronically increased stimulation and subsequent PKA activation could result in a robust apoptotic response.6 Moreover, it was shown that the pro-apoptotic BH3-only protein Bim has a crucial role in cAMP-induced apoptosis in the T cell hybridoma line S49.21 Therefore, our initial task was to explore the generality of this phenomenon in different tissues. We could demonstrate PKA activation and subsequent Bim induction both at the protein and mRNA levels by a variety of stimuli and in a variety of tissues (Figures 1 and 4). We could also demonstrate that in thymic and heart tissues, Bim has a critical role in βAR-mediated apoptosis (Figure 2). Our results also demonstrate that Bim has an important role in the heart development. In MEFs and thymocytes, there was a lag time between mRNA induction and protein accumulation. This could be partly due to the Bim protein stabilization by PKA, as we previously reported.17 Lag time between transcript induction and protein accumulation has been reported in many previous studies,45 particularly for the βAR pathway where feedback inhibition and receptor decoupling leads to receptor desensitization.46
Apart from protecting from βAR-induced cardiomyopathy, Bim−/− mice had a significantly larger heart (Supplementary Figure S2d). This is not associated with hyperplasia rather because of increased size of cardiomyocytes, which does not change with isoproterenol treatment (Figures 3a and b). As cardiac sympathetic nervous system is a strong regulator of cardiomyocyte size via βAR-dependent repression of proteolysis,47 we believe that this size increase is a function of elevated βAR activity in the absence of Bim, similar to the observation in thymocytes. Furthermore, this hypertrophy is not associated with any pathology, as we could not detect any upregulation of pathology-associated genes such as Myh6 and 7; Serca2 or Col1a1 and Col3a1.48 However, cardiac skeletal α-actin (Aska) was significantly increased in the KO tissue. On the basis of gene knock-out studies, it is believed that Aska contributes to muscular strength and contractility,49 and thus our results suggest that blocking Bim expression in the heart will maintain the βAR-mediated inotropic function and only prevent the apoptotic arm of the signal transduction pathway.
The observation that deletion of Prkar1a results in Bim-dependent apoptosis is significant in yet another context (Figure 4). Inactivation of Prkar1a has been attributed to the development of cancer, such as multiple endocrinal neoplasia, myxoma and thyroid tumors.50 Therefore, Prkar1a is believed to be a tumor suppressor.50 However, our results suggest that cancer associated with Prkar1a mutations can develop only if there are cooperating oncogenic mutations that inhibit Bim-induced apoptosis. This could involve either genetic or epigenetic changes in the Bim gene or mutations that counter Bim induction, such as overexpression of antiapoptotic Bcl-2 family members. Given that Bim is a tumor suppressor in many cancers,41, 51 it is possible that in those cancers where Prkar1a loss of heterozygosity causes elevated PKA activity, Bim is a critical tumor suppressor.
Our data show for the first time that c-Myc regulates the transcriptional induction of Bim during βAR activation. CREB is one of the main downstream targets of PKA and based on expression analysis, it was reported that Bim was a downstream target of CREB.52 However, our results, obtained through a variety of methods, demonstrate that it is c-Myc that is mediating Bim transcriptional induction (Figure 5). This finding is also relevant to Eμ-myc transgenic mice, a model of Burkitt’s lymphoma, where deregulated c-Myc expression leads to increased Bim protein levels in B cells,36 and thereby suppresses lymphoma development. However, c-Myc was considered to upregulate Bim indirectly. The results presented here represent the first demonstration of a direct role for c-Myc in Bim transcriptional upregulation.
Yet another significant finding in this study is the role of CBP in Bim induction. Our results demonstrated an absolute requirement of this transcriptional co-factor for βAR stimulation-triggered Bim induction (Figure 6). Mutations in CBP are found in B cell lymphomas.40 It is therefore possible that Bim downregulation in these cells may contribute to tumorigenesis, given that Bim functions as a tumour suppressor in B cell lymphoma.36 Our results also show that although LK-2 cells have normal expression of p300, Bim can be upregulated only when CBP is forcibly expressed (Figure 6e and Supplementary Figures S4c and d). This indicates that there is no functional redundancy between Cbp and p300 in inducing Bim transcription in spite of the extensive sequence similarity between these two proteins, in agreement with previous report.53 Finally, our findings of epigenetic changes at the Bim locus (Figure 7), both in terms of histone acetylation and DNA methylation during PKA activation/CBP expression can be reconciled as CBP is a HAT and the interplay between these two processes in gene regulation has long been established.54 On the basis of these findings, the following model (Figure 7c) is proposed where βAR-mediated PKA activation leads to activation of CBP. The activated CBP leads to histone acetylation (and demethylation) at the Bim promoter locus, leading to increased gene expression.
Taken together, these results provide a clear link between cAMP/PKA pathway activation and Bim induction and consequent Bim-mediated apoptosis. Catecholamines are the prime mediators of cardiomyocyte apoptosis.6 In the immune system, apart from the sympathetic nervous innervation of lymphoid organs, lymphocytes and macrophages themselves can be a source of catecholamines.4, 24 This is known to contribute to thymocyte apoptosis and thymic involution can exert immune-modulatory effect as a prototypic stress response.24 The present work is the first demonstration of the molecular pathway of βAR-mediated apoptosis. Understanding the molecular mechanism of this apoptotic pathway may provide critical clues for developing new drugs for the treatment of diseases associated with increased β-adrenergic activation.
Materials and Methods
Cell culture and luciferase assay
Cells were grown in DMEM supplemented with 10% foetal calf serum at 10% CO2 and at 37 °C. Transient transfections were performed using Fugene (Roche, Indianapois, IN, USA). For PKACα or c-Myc-expressing cells, proteins expression was induced 24 h after transfection with 5 nm 4-OHT overnight, followed by luciferase assay using the Dual Luciferase (Promega, Alexandria, NSW, Australia) kit. For catecholamine induction, cells were treated 24 h after transfection, with isoproterenol (10 μM) and rolipram (10 μM) in serum-free medium overnight before the luciferase assay.
Animal experimentation
All animal experiments were conducted according to the La Trobe University Animal Ethics committee and the Alfred Medical Research and Education Precinct Animal Ethics Committee guidelines.
Lentiviral infection
To generate stable, transformed MEFs, primary MEFs were infected with lentiviral particles expressing SV40 T antigen. To generate conditional knock-out MEFs, transformed MEFs (Prkar1afl/fl and c-Mycfl/fl) were infected with 4-hydroxytamoxifen (4-OHT)-inducible CRE-expressing lentiviral particles and clones were selected. CRE deletion was induced by the addition of 4-OHT (5 nM). Production of lentiviral particles and infections were carried out as described.17
Foetal thymic organ culture
Thymic organ culture was performed as described.55 Thymus lobes obtained from E15 embryos were cultured in DMEM medium supplemented with 50 μM β-mercaptoethanol (Sigma, Castle Hill, NSW, Australia) plus 10% FCS. After 12 days, thymocytes were treated with 10 μM isoproterenol and 0.5 mM IBMX. Cells were isolated from these thymi for FACS, qPCR, western blot and cAMP analyses.
RT-quantitative PCR
Total RNA from thymocytes was isolated using TRIZOL. Complementary DNA (cDNA) was synthesized from 2 μg of total RNA using the Superscript III RT-PCR system (Catalogue number 18080-051 Invitrogen, Carlsbad, CA, USA), using oligo-dT primer, according to manufacturer’s instructions. qPCR was carried out using Brilliant II SYBER Green QPCR master Mix (Catalogue number 600828 Stratagene, La Jolla, CA, USA). All cDNA samples were tested in triplicate using a Light cycler 480 Real-time PCR instrument (Roche). Following primers were used for qPCR reactions: Bim (F): 5′-GAGTTGTGACAAGTCAACACAAACC-3′; Bim (R): 5′-GAAGATAAAGCGTAACAGTTGTAAGATA-3′; Nur77 (F): 5′-CCTGTTGCTAGAGTCTGCCTTC-3′; Nur77 (R): 5′-CAATCCAATCACCAAAGCCACG-3′; Puma (F): 5′-ATGCCTGCCTCACCTTCATCT-3′; Puma (R): 5′-AGCACAGGATTCACAGTCTGGA-3′; Noxa (F): 5′-ACTGTGGTTCTGGCGCAGAT-3′; Noxa (R): 5′-TTGAGCACACTCGTCCTTCAA-3′; c-Myc (F): 5′-CAAATCCTGTACCTCGTCCGATTC-3′; c-Myc (R): 5′-CTTCTTGCTCTTCTTCAGAGTCGC-3′; Serca2a (F): 5′-TCGACCAGTCAATTCTTACAG-3′; Serca2a (R): 5′-GGGACAGGGTCAGTATGCTT-3′; Myh6 (F): 5′-GTCACCAACAACCCATACGACTAC-3′; Myh6 (R): 5′-CAGCACATCAAAGGCACTATCAGT-3′; Myh7 (F): 5′-AGAACCTACTGCGGCTCCA-3′; Myh7 (R): 5′-CTACTCCTCATTCAGGCC-3′; Col1a1 (F): 5′-GGAGATGATGGGGAAGCTG-3′; Col1a1 (R): 5′-AATCCACGAGCACCCTGA-3′; Col3a1 (F): 5′-GGAATGGAGCAAGACAGTCTTTG-3′; Col3a1 (R): 5′-TGCGATATCTATGATGGGTAGTCTCA-3′; Aska (F): 5′-CCTGCCATGTATGTGGCTATC-3′; Aska (R): 5′-CCCCAGAATCCAACACGAT-3′; 18s rRNA (F): 5′-CCGCTCCCAAGATCCAACTA-3′; 18s rRNA (R): 5′-TTGGAGGGCAAGTCTGGTG-3′.
Chromatin immunoprecipitations
DNA for ChIP analysis was prepared from cells expressing PKACα under 4-OHT regulation using EZ-ChIP (Millipore, Billerica, MA, USA) kit, following manufacturer’s instructions. The following antibodies were used: anti-Myc (N-262, Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-CBP (C-20, Santa Cruz Biotechnology), anti-acetyl-Histone H3 (17-658, Millipore), anti-acetyl-Histone H4 (17-630, Millipore). Immunoprecipitated DNAs were subjected to PCR using oligos specific to Bim promoter F: 5′-GTGAAGTGCTAACTAGATTGCAC-3′ and R: 5′-GAAAACCAGCAGTGGTGGAC-3′. Primers specific to the 3′UTR were used as negative controls, that is, F: 5′-TGTCTTCCTGCTGATTCAGC-3′ and R: 5′-AAACGTAAAGGAAGCCAGGG-3′.
Western and northern blot analyses
Western blot and northern blot analyses were performed as described before.17 The following antibodies were used for western blot analyses: anti-Bim (3C5), anti-HA (6E2, Cell Signaling, Boston, MA, USA), anti-CBP, anti-p300 (N-15, Santa Cruz Biotechnology), anti pCREB Ser133, Affinity Bioreagents, Golden, CO, USA), anti HSP70, anti-acetyl lysine (9441, Cell Signaling), anti PRKAR1A (610609, BD Biosciences, CA, USA), anti-PKA substrate (9621, Cell Signaling), anti-PUMA (3043, ProSci, Poway, CA, USA).
cAMP measurement
cAMP in cellular extracts was measured using cAMP Direct Immunoassay Kit (Cat# ab65355, Abcam, Cambridge, MA, USA).
Cardiomyocyte isolation
Cardiomyocytes were isolated by Langendorff perfusion technique, as described.56
In vitro PKA kinase assay
In vitro PKA kinase assay was performed as described.17
Catecholamine measurements
Blood samples were collected from anesthetized animals by cardiac puncture. Plasma were prepared by centrifugation and stored at −80 °C, until assay with high-performance liquid chromatography (HPLC).57 This method allowed simultaneous determination of norepinephrine and epinephrine. Catecholamines were adsorbed with activated alumina, separated by HPLC, and quantified by electrochemical detection.
Histology
Hearts were fixed in 10% formalin in phosphate-buffered saline, paraffin-embedded, serially sectioned (5 μM) and stained with Picrosirius red or Masson’s trichrome.58
Echocardiography
Animals were anesthetized by isoflurane. Using an IE33 ultrasound system and a linear 15 MHz probe, two-dimentional (2D) short-axis loop of the LV was acquired for measurement of LV cross-sectional areas at the diastole and systole. LV fractional shortening was calculated from LV diameters at the diastole and systole from the M-mode traces derived from 2D image, as described before.58
Promoter methylation analysis
Promoter methylation was analysed by bisulphite sequencing of the BIM promoter using MethylDetector kit (Active Motif, Carlsbad, CA, USA). Sulphonated DNA samples (from Ramos, LK-2 and LK-2 cells expressing functional CBP) were amplified by PCR using primers specific for CpG island corresponding to bases −1513 to −948 (relative to the transcription start site on human BIM gene). PCR-amplified fragments were cloned into PCRII-TOPO vector using TOPO TA Cloning kit (Life Technologies, Grand Island, NY, USA). Ten clones from each cloning were sequenced and analysed using BISMA software (Jacobs University Breman, Germany).
Abbreviations
- cAMP/PKA:
-
cyclic AMP-regulated protein kinase A
- βAR:
-
β-adrenergic receptor
- CREB:
-
cAMP response element-binding protein
- CBP:
-
CREB-binding protein
- 4-OHT:
-
4-hydroxytamoxifen
- FTOC:
-
foetal thymic organ culture
- MHC:
-
myosin heavy chain
- Prkar1a:
-
PKA regulatory subunit 1a
- ChIP:
-
chromatin immunoprecipitation
- EMSA:
-
electrophoretic mobility shift assay
References
Sutherland EW . On the biological role of cyclic AMP. Jama 1970; 214: 1281–1288.
Tomkins GM . The metabolic code. Science 1975; 189: 760–763.
Pratt RM, Martin GR . Epithelial cell death and cyclic AMP increase during palatal development. Proc Natl Acad Sci USA 1975; 72: 874–877.
Flierl MA, Rittirsch D, Nadeau BA, Chen AJ, Sarma JV, Zetoune FS et al. Phagocyte-derived catecholamines enhance acute inflammatory injury. Nature 2007; 449: 721–725.
Stavrakis S, Yu X, Patterson E, Huang S, Hamlett SR, Chalmers L et al. Activating autoantibodies to the beta-1 adrenergic and m2 muscarinic receptors facilitate atrial fibrillation in patients with Graves' hyperthyroidism. J Am Coll Cardiol 2009; 54: 1309–1316.
Xiao RP . Beta-adrenergic signaling in the heart: dual coupling of the beta2-adrenergic receptor to G(s) and G(i) proteins. Sci STKE 2001; 16.
Annane D, Bellissant E, Cavaillon J-M . Septic shock. Lancet 2005; 365: 63–78.
Oberbeck R, Schmitz D, Wilsenack K, Schuler M, Pehle B, Schedlowski M et al. Adrenergic modulation of survival and cellular immune functions during polymicrobial sepsis. Neuroimmunomodulation 2004; 11: 214–223.
Shinkai T, Zhang L, Mathias SA, Roth GS . Dopamine induces apoptosis in cultured rat striatal neurons; possible mechanism of D2-dopamine receptor neuron loss during aging. J Neurosci Res 1997; 47: 393–399.
Padgett DA, Glaser R . How stress influences the immune response. Trends Immunol 2003; 24: 444–448.
Bouillet P, Metcalf D, Huang DC, Tarlinton DM, Kay TW, Kontgen F et al. Proapoptotic Bcl-2 relative Bim required for certain apoptotic responses, leukocyte homeostasis, and to preclude autoimmunity. Science 1999; 286: 1735–1738.
Puthalakath H, O’Reilly LA, Gunn P, Lee L, Kelly PN, Huntington ND et al. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007; 129: 1337–1349.
Bouillet P, Purton JF, Godfrey DI, Zhang LC, Coultas L, Puthalakath H et al. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature 2002; 415: 922–926.
Enders ABP, Puthalakath H, Xu Y, Tarlinton DM, Strasser A . Loss of the pro-apoptotic BH3-only Bcl-2 family member Bim inhibits BCR stimulation-induced apoptosis and deletion of autoreactive B cells. J Exp Med 2003; 198: 1119–1126.
Kuroda J, Puthalakath H, Cragg MS, Kelly PN, Bouillet P, Huang DC et al. Bim and Bad mediate imatinib-induced killing of Bcr/Abl+ leukemic cells, and resistance due to their loss is overcome by a BH3 mimetic. Proc Natl Acad Sci USA 2006; 103: 14907–14912.
Cragg MS, Kuroda J, Puthalakath H, Huang DC, Strasser A . Gefitinib-induced killing of NSCLC cell lines expressing mutant EGFR requires BIM and can be enhanced by BH3 mimetics. PLoS Med 2007; 4: 1681–1689 discussion 1690.
Moujalled D, Weston R, Anderton H, Ninnis R, Goel P, Coley A et al. Cyclic-AMP-dependent protein kinase A regulates apoptosis by stabilizing the BH3-only protein Bim. EMBO Rep 2011; 12: 77–83.
Singh K, Communal C, Sawyer DB, Colucci WS . Adrenergic regulation of myocardial apoptosis. Cardiovasc Res 2000; 45: 713–719.
Leposavic G, Ugresic N, Pejcic-Karapetrovic B, Micic M . Castration of sexually immature rats affects sympathetic innervation of the adult thymus. Neuroimmunomodulation 2000; 7: 59–67.
Gruver AL, Sempowski GD . Cytokines, leptin, and stress-induced thymic atrophy. J Leukoc Biol 2008; 84: 915–923.
Zhang LIP . The pro-apoptotic protein Bim is a convergence point for cAMP/protein kinase A- and glucocorticoid-promoted apoptosis of lymphoid cells. J Biol Chem 2004; 279: 20858–20865.
Kim S, Bakre M, Yin H, Varner JA . Inhibition of endothelial cell survival and angiogenesis by protein kinase A. J Clin Invest 2002; 110: 933–941.
Anagnostou VK, Doussis-Anagnostopoulou I, Tiniakos DG, Karandrea D, Agapitos E, Karakitsos P et al. Ontogeny of intrinsic innervation in the human thymus and spleen. J Histochem Cytochem 2007; 55: 813–820.
Toti P, De Felice C, Stumpo M, Schurfeld K, Di Leo L, Vatti R et al. Acute thymic involution in fetuses and neonates with chorioamnionitis. Hum Pathol 2000; 31: 1121–1128.
Maruoka H, Sasaya H, Shimamura Y, Nakatani Y, Shimoke K, Ikeuchi T . Dibutyryl-cAMP up-regulates nur77 expression via histone modification during neurite outgrowth in PC12 cells. J Biochem 2010; 148: 93–101.
Shizukuda Y, Buttrick PM . Subtype specific roles of beta-adrenergic receptors in apoptosis of adult rat ventricular myocytes. J Mol Cell Cardiol 2002; 34: 823–831.
Zhu WZ, Wang SQ, Chakir K, Yang D, Zhang T, Brown JH et al. Linkage of beta1-adrenergic stimulation to apoptotic heart cell death through protein kinase A-independent activation of Ca2+/calmodulin kinase II. J Clin Invest 2003; 111: 617–625.
Du XJ, Gao XM, Wang B, Jennings GL, Woodcock EA, Dart AM . Age-dependent cardiomyopathy and heart failure phenotype in mice overexpressing beta(2)-adrenergic receptors in the heart. Cardiovasc Res 2000; 48: 448–454.
Woodcock EA, Du XJ, Reichelt ME, Graham RM . Cardiac alpha 1-adrenergic drive in pathological remodelling. Cardiovasc Res 2008; 77: 452–462.
Josefsson E, Bergquist J, Ekman R, Tarkowski A . Catecholamines are synthesized by mouse lymphocytes and regulate function of these cells by induction of apoptosis. Immunology 1996; 88: 140–146.
Du XJ, Cole TJ, Tenis N, Gao XM, Kontgen F, Kemp BE et al. Impaired cardiac contractility response to hemodynamic stress in S100A1-deficient mice. Mol Cell Biol 2002; 22: 2821–2829.
Morel E, Marcantoni A, Gastineau M, Birkedal R, Rochais F, Garnier A et al. cAMP-binding protein Epac induces cardiomyocyte hypertrophy. Circ Res 2005; 97: 1296–1304.
Pasqualucci LKY, Gu H, Dalla-Favera R . PKA-mediated phosphorylation regulates the function of activation-induced deaminase (AID) in B cells. Proc Natl Acad Sci USA 2006; 103: 395–400.
Gangoda L, Doerflinger M, Lee YY, Rahimi A, Etemadi N, Chau D et al. Cre transgene results in global attenuation of the cAMP/PKA pathway. Cell Death Dis 2012; 3: e365.
Kim J, Lee JH, Iyer VR . Global identification of Myc target genes reveals its direct role in mitochondrial biogenesis and its E-box usage in vivo. PLoS ONE 2008; 3.
Egle A, Harris AW, Bouillet P, Cory S . Bim is a suppressor of Myc-induced mouse B cell leukemia. Proc Natl Acad Sci USA 2004; 101: 6164–6169.
Yin X, Giap C, Lazo JS, Prochownik EV . Low molecular weight inhibitors of Myc-Max interaction and function. Oncogene 2003; 22: 6151–6159.
Wu KJMM, Morse HC 3rd, Dalla-Favera R . c-MYC activates protein kinase A (PKA) by direct transcriptional activation of the PKA catalytic subunit beta (PKA-Cbeta) gene. Oncogene 2002; 21: 7872–7882.
Vervoorts J, Luscher-Firzlaff JM, Rottmann S, Lilischkis R, Walsemann G, Dohmann K et al. Stimulation of c-MYC transcriptional activity and acetylation by recruitment of the cofactor CBP. EMBO Rep 2003; 4: 484–490.
Pasqualucci L, Dominguez-Sola D, Chiarenza A, Fabbri G, Grunn A, Trifonov V et al. Inactivating mutations of acetyltransferase genes in B-cell lymphoma. Nature 2011; 471: 189–195.
Tagawa H, Karnan S, Suzuki R, Matsuo K, Zhang X, Ota A et al. Genome-wide array-based CGH for mantle cell lymphoma: identification of homozygous deletions of the proapoptotic gene BIM. Oncogene 2005; 24: 1348–1358.
Kishimoto M, Kohno T, Okudela K, Otsuka A, Sasaki H, Tanabe C et al. Mutations and deletions of the CBP gene in human lung cancer. Clin Cancer Res 2005; 11 (2 Pt 1): 512–519.
De Bruyne E, Bos TJ, Schuit F, Van Valckenborgh E, Menu E, Thorrez L et al. IGF-1 suppresses Bim expression in multiple myeloma via epigenetic and posttranslational mechanisms. Blood 2010; 115: 2430–2440.
Mancini M, Veljkovic N, Leo E, Aluigi M, Borsi E, Galloni C et al. Cytoplasmatic compartmentalization by Bcr-Abl promotes TET2 loss-of-function in chronic myeloid leukemia. J Cell Biochem 2012; 113: 2765–2774.
McAdams HH, Arkin A . Stochastic mechanisms in gene expression. Proc Natl Acad Sci USA 1997; 94: 814–819.
Seibold A, January BG, Friedman J, Hipkin RW, Clark RB . Desensitization of beta2-adrenergic receptors with mutations of the proposed G protein-coupled receptor kinase phosphorylation sites. J Biol Chem 1998; 273: 7637–7642.
Zaglia T, Milan G, Franzoso M, Bertaggia E, Pianca N, Piasentini E et al. Cardiac sympathetic neurons provide trophic signal to the heart via beta2- adrenoceptor dependent regulation of proteolysis. Cardiovas Res 2012; 97: 240–250.
Zwadlo C, Borlak J . Disease-associated changes in the expression of ion channels, ion receptors, ion exchangers and Ca(2+)-handling proteins in heart hypertrophy. Toxicol Appl Pharmacol 2005; 207: 244–256.
Crawford K, Flick R, Close L, Shelly D, Paul R, Bove K et al. Mice lacking skeletal muscle actin show reduced muscle strength and growth deficits and die during the neonatal period. Mol Cell Biol 2002; 22: 5887–5896.
Nadella KS, Jones GN, Trimboli A, Stratakis CA, Leone G, Kirschner LS . Targeted deletion of Prkar1a reveals a role for protein kinase A in mesenchymal-to-epithelial transition. Cancer Res 2008; 68: 2671–2677.
Zantl N, Weirich G, Zall H, Seiffert BM, Fischer SF, Kirschnek S et al. Frequent loss of expression of the pro-apoptotic protein Bim in renal cell carcinoma: evidence for contribution to apoptosis resistance. Oncogene 2007; 26: 7038–7048.
Huseby S, Gausdal G, Keen TJ, Kjaerland E, Krakstad C, Myhren L et al. Cyclic AMP induces IPC leukemia cell apoptosis via CRE-and CDK-dependent Bim transcription. Cell Death Dis 2011; 2: e237.
Vo N, Goodman RH . CREB-binding protein and p300 in transcriptional regulation. J Biol Chem 2001; 276: 13505–13508.
Dobosy JR, Selker EU . Emerging connections between DNA methylation and histone acetylation. Cell Mol Life Sci 2001; 58: 721–727.
Ramsdell F, Zuniga-Pflucker JC, Takahama Y . In vitro systems for the study of T cell development: fetal thymus organ culture and OP9-DL1 cell coculture. Curr Protoc Immunol 2006; 3: 18.
Wolska BM, Solaro RJ . Method for isolation of adult mouse cardiac myocytes for studies of contraction and microfluorimetry. Am J Physiol 1996; 271 (3 Pt 2): H1250–H1255.
Du XJ, Esler MD, Dart AM . Sympatholytic action of intravenous amiodarone in the rat heart. Circulation 1995; 91: 462–470.
Xu Q, Dalic A, Fang L, Kiriazis H, Ritchie RH, Sim K et al. Myocardial oxidative stress contributes to transgenic beta-adrenoceptor activation-induced cardiomyopathy and heart failure. Br J Pharmacol 2011; 162: 1012–1028.
Acknowledgements
We thank Vishy Iyer, Andreas Strasser, Liam O’Connor, Daniel Gray, Holly Anderton, Diep Chau, Lorraine O’Reilly, Hua Gu and Warren Alexander for reagents and advice. YYL and LG are supported by CRC BT, X-JD is supported by NHMRC project grant (APP1005329) and HP is supported by ARC Future Fellowship (FT0990683) and by ARC project grant (DP110100417).
Author Contributions
HP, X-J D., PB and Id A designed the experiments. YYL, MD, LG, RW, AR, DM, MH, X-JD and HP performed the experiments and YYL, X-JD and HP analysed the data, and HP prepared the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Edited by M Piacentini
Supplementary Information accompanies this paper on Cell Death and Differentiation website
Rights and permissions
About this article
Cite this article
Lee, Y., Moujalled, D., Doerflinger, M. et al. CREB-binding protein (CBP) regulates β-adrenoceptor (β-AR)−mediated apoptosis. Cell Death Differ 20, 941–952 (2013). https://doi.org/10.1038/cdd.2013.29
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cdd.2013.29
Keywords
This article is cited by
-
Bim Deletion Reduces Functional Deficits Following Ischemic Stroke in Association with Modulation of Apoptosis and Inflammation
NeuroMolecular Medicine (2022)
-
Mechanistic insight into sevoflurane-associated developmental neurotoxicity
Cell Biology and Toxicology (2022)
-
Using antagonistic pleiotropy to design a chemotherapy-induced evolutionary trap to target drug resistance in cancer
Nature Genetics (2020)
-
A siRNA screen reveals the prosurvival effect of protein kinase A activation in conditions of unresolved endoplasmic reticulum stress
Cell Death & Differentiation (2016)
-
Loss of Prkar1a leads to Bcl-2 family protein induction and cachexia in mice
Cell Death & Differentiation (2014)

{kind=link}
{kind=link}
{kind=link}
{kind=link}