
Abstract
Iron is a catalyst in the formation of reactive oxygen species. Oxidative stress is associated with the pathogenesis of both human and experimental animal models of renovascular hypertension. We hypothesized that iron is involved in the pathogenesis of renovascular hypertension and that iron restriction may affect the pathogenesis of renovascular hypertension via the inhibition of oxidative stress. Herein, we investigated the effect of iron restriction on hypertension and renal damage in a rat model of two-kidney one-clip (2K1C) renovascular hypertension. Renovascular hypertension was induced by 2K1C in male Sprague–Dawley rats. At the day of clipping, 2K1C rats were divided into untreated (2K1C) and dietary iron-restricted groups (2K1C+IR). The 2K1C rats showed hypertension after the day of clipping, whereas dietary iron restriction attenuated the development of hypertension. Vascular hypertrophy and the increased fibrotic area were suppressed in the 2K1C+IR group. The clipped kidney developed renal atrophy in both the 2K1C and 2K1C+IR groups after clipping. However, the unclipped kidney showed renal hypertrophy in the 2K1C and 2K1C+IR groups, and the extent was less in the 2K1C+IR group. The 2K1C rats exhibited glomerulosclerosis and tubulointerstitial fibrosis in the unclipped kidney, whereas these changes were attenuated by an iron-restricted diet. Importantly, proteinuria was decreased in the 2K1C+IR group, along with decreased urinary 8-hydroxy-2'-deoxyguanosine excretion and superoxide production of the unclipped kidney. Moreover, the expression of nuclear mineralocorticoid receptor in the unclipped kidney of the 2K1C rats was attenuated by iron restriction. These data indicate a novel effect of iron restriction on hypertension and renal damage in renovascular hypertension.
Similar content being viewed by others

Introduction
Secondary hypertension is often resistant to standard antihypertensive treatment. Renovascular hypertension (RVHT) is the most common type of secondary hypertension, accounting for 1 to 5% of patients with hypertension.1 RVHT is caused by stenosis or obstruction of the renal artery, and the etiology of RVHT is mostly atherosclerosis in elderly patients and fibromuscular dysplasia in younger patients. Stenosis of the renal artery may result in both hypertension and chronic kidney disease.2 The management of RVHT includes antihypertensive medications, percutaneous transluminal renal angioplasty and surgery. In patients with atherosclerotic RVHT, randomized clinical trials have not shown any advantage of percutaneous transluminal renal angioplasty over antihypertensive medications in terms of hypertension or chronic kidney disease controls.3, 4 Thus, the management for RVHT remains controversial.
Iron is a vital element necessary for life. However, iron is a catalyst in the formation of reactive oxygen species, which induces cell and tissue damage. Therefore, iron is considered to be associated with the pathogenesis of several cardiovascular and renal diseases. In both human and experimental animal models of RVHT, increased oxidative stress is associated with the pathogenesis of RVHT;5, 6 however, there has been no report assessing the pathogenesis of RVHT in terms of iron so far. Thus, we hypothesized that iron is involved in the pathogenesis of RVHT and that iron restriction (IR) may affect the pathogenesis of RVHT via the inhibition of oxidative stress.
The two-kidney one-clip (2K1C) hypertensive rat is an experimental model that, in many respects, resembles human RVHT.7 In the present study, we assessed the effect of dietary IR on hypertension and renal damage in 2K1C rats. Herein, we demonstrate that IR attenuates the development of hypertension and renal damage in 2K1C rats.
Methods
Rat model of RVHT
Five-week-old male Sprague–Dawley rats were fed a normal diet for 1 week. Thereafter, RVHT was induced using the Goldblatt 2K1C method, as described previously.8, 9 A right retroperitoneal flank incision was performed. Next, the right renal artery was exposed and occluded with a U-shaped silver clip with an internal diameter of 0.20 mm on the vessel.
Protocol 1
At the day of clipping, 2K1C rats were randomly divided into two groups and were given a normal diet (RVHT, n=10) or an iron-restricted diet (RVHT+IR, n=10) for 8 weeks. Sham-operated animals on a normal diet served as a control (Control, n=10). The nutrients of the normal diet consisted of 33% cornstarch, 22% casein, 5% cellulose, 30% sucrose, 5% corn oil, 4% of a mineral mixture and 1% of a vitamin mix. The mineral mixture contains 0.43% CaHPO4·2H2O, 34.31% KH2PO4, 25.06% NaCl, 0.623% FeC6H5O7·5H2O, 4.8764% MgSO4, 0.02% ZnCl2, 0.121% MnSO4·5H2O, 0.156% CuSO4·5H2O, 0.0005% KI, 29.29% CaCO3, 0.0025% (NH4)6MO7O24·4H2O and 5.11% microcrystalline cellulose. The iron-restricted diet was based on the normal diet but with a mineral mixture free of FeC6H5O7·5H2O.
Protocol 2
At the day of clipping, 2K1C rats were randomly assigned to the two groups and were given a normal diet (RVHT, n=6) or a 30% iron-restricted diet (RVHT+mild IR, n=6) for 8 weeks. Sham-operated animals on a normal diet served as a control (Control, n=6). The nutrients of the normal diet were same as that of protocol 1. A 30% iron-restricted diet was based on the normal diet with a mineral mixture of 0.1869% FeC6H5O7·5H2O. The rats were maintained on a 12 h light/dark cycle and had free access to food and water. All of our experimental procedures were approved by the Animal Research Committee of Hyogo College of Medicine. At 8 weeks after surgery, the rats were killed. Their blood was quickly withdrawn by abdominal aorta puncture, and the serum and plasma were stored at −80 °C before analysis. The tissues were resected and washed in phosphate-buffered saline. Thereafter, the organs were quickly snap frozen in liquid nitrogen and stored at −80 °C.
Assessments of blood pressure, heart rate, urine, blood and tissue iron content
Systolic blood pressure and heart rate were measured using a noninvasive computerized tail-cuff system in conscious rats every week after surgery (MK-2000; Muromachi Kikai, Tokyo, Japan).10 Twenty-four-hour urine samples were collected in metabolic cages at 7 weeks after surgery. The urinary concentrations of total protein, sodium and potassium were determined using the pyrogallol red method and electrode method, respectively.11 The urinary concentrations of 8-hydroxy-2'-deoxyguanosine (8-OHdG) and iron were determined by enzyme-linked immune sorbent assay (Japan Institute for the Control of Aging, Shizuoka, Japan) and the atomic absorption method, respectively. The peripheral blood cell count was measured using an automatic cell count analyzer (Pentra 60 LC-5000; Horiba, Kyoto, Japan). Serum iron and erythropoietin levels were determined as reported previously.12 The plasma aldosterone level was measured by the radioimmunoassay (SPAC-S Aldosterone Kit; TFB, Tokyo, Japan). The renal iron content was measured using the Metallo Assay Kit according to the manufacturer’s instructions (AKJ Global Technology, Chiba, Japan). The iron content was then corrected to the kidney weight for each sample.13
Gene expression analysis
RNA was extracted from the kidney using TRIzol reagent (Invitrogen). DNase-treated RNA was reverse transcribed into cDNA using random primers (Applied Biosystems). Real-time PCR assays were performed using the ABI PRISM 7900 with TaqMan Universal PCR Master Mix and TaqMan Gene Expression Assays (Applied Biosystems).12 The TaqMan Gene Expression Assays used as primers and probes for each gene were as follows: collagen type I (assay ID Rn00801649_g1), collagen type III (assay ID Rn01437683_m1), CD68 (assay ID Rn01495634_g1), monocyte chemoattractant protein-1 (MCP-1; assay ID Rn00580555_m1), serum and glucocorticoid regulated kinase 1 (SGK1; assay ID Rn00570285_m1) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; assay ID Rn99999916_s1). GAPDH was used as an internal control.
Western blot analysis
Aortic tissues were homogenized with ice-cold lysis buffer as described previously,10 and nuclear extracts from the kidney were isolated with commercially available kits (BioVision, Livingston, NJ, USA). The protein homogenate from the aorta and nuclear extracts from the kidney were separated by SDS-PAGE and transferred onto polyvinylidene difluoride membranes. The expression levels were detected using an Enhanced Chemiluminescence Kit (Thermo Scientific, Rockford, IL, USA). Here, the primary antibodies used were rabbit anti-phospho-endothelial nitric oxide synthase (Ser1177) (Cell Signaling Technology, Danvers, MA, USA; 1:1000 dilution), rabbit anti-endothelial nitric oxide synthase (Santa Cruz, Dallas, TX, USA; 1:1000 dilution), rabbit anti-mineralocorticoid receptor (MR) (Perseus Proteomics, Tokyo, Japan; 1:1000 dilution) and rabbit anti-cAMP response element binding protein (Millipore, Billerica, MA, USA; 1:1000 dilution).
Histological analysis
The aorta and kidney tissues were fixed with buffered 4% paraformaldehyde, embedded in paraffin and cut into 4-μm-thick sections. Hematoxylin–eosin staining of the aorta, periodic acid-Schiff staining of the kidney and Masson’s trichrome staining of the aorta and kidney were performed using serial sections. Aortic fibrotic lesions were evaluated by the semiquantitative score using the ImageJ software (National Institute of Health, Bethesda, MD, USA). Glomerular and tubular lesions were evaluated using the semiquantitative score method as described previously.12 The clipped kidney sections were stained using the von Kossa method (Polysciences, Warrington, PA, USA). Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining was performed according to the manufacturer’s protocol (TaKaRa, Shiga, Japan). The section was counterstained with DAPI (4',6-diamidino-2-phenylindole). The numbers of TUNEL-positive cells per 60 glomeruli and tubules in 10 randomly selected fields were counted. Kidney tissues were also quickly embedded in Tissue-Tek OCT compound (Sakura Finetechnical, Tokyo, Japan) and snap frozen in liquid nitrogen. Superoxide detection was performed on transverse cross-sections of 8 μm thickness incubated with dihydroethidium as described previously.10 The dihydroethidium fluorescence intensity was measured by the semiquantitative score using the ImageJ software.
Statistical analysis
Values are reported as the means±s.e.m. Statistical analysis was performed using one-way analysis of variance. Analysis of variance (Kruskal–Wallis test, followed by Mann–Whitney U-test) was used for statistical comparisons. Probability values <0.05 were considered to be significant.
Results
Impact of IR on the development of hypertension in renovascular hypertensive rats
Systolic blood pressure increased after the day of clipping and was reduced gradually after 3 weeks of clipping in the 2K1C group (Figure 1a). By contrast, dietary IR attenuated the initial development of hypertension from the day of clipping in the 2K1C rats (Figure 1a). The heart rate was not significantly different among the groups throughout the experimental period (Figure 1b). The phosphorylation of endothelial nitric oxide synthase at Ser1177 in the aorta was not different among the groups at 8 weeks after clipping (Figure 1c); however, as shown in Figure 1d, vascular hypertrophy and an increased fibrotic area were observed in the 2K1C group compared with the control group at 8 weeks after clipping, and these pathologic changes were suppressed in the 2K1C+IR group (Figures 1d and e). Body weight was lower in the 2K1C+IR group compared with that in the other groups (Table 1). Anemia was observed in the 2K1C+IR groups at 8 weeks after clipping. The serum iron levels were decreased and serum erythropoietin levels were increased in the 2K1C+IR group. During the experimental period, there was no significant difference in the feeding behavior and activity among the groups, and none of the rats died in any of the groups.

Impact of iron restriction (IR) on hypertension and renal structure of the clipped kidney in two-kidney one-clip (2K1C) rats. Time course of (a) systolic blood pressure and (b) heart rate in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). (c) Aortic phosphorylated (top) and total (bottom) expression of endothelial nitric oxide synthase (eNOS) in the Control, 2K1C and 2K1C+IR groups (n=3 in each group). (d) Representative images of hematoxylin–eosin (HE) and Masson’s trichrome (MT) staining of the aorta sections. Scale bars, 100 μm. (e) Quantitative analysis of aortic wall thickness and fibrosis area in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). (f) Representative gross morphology of the clipped kidney and clipped kidney weight/TL ratio in the Control, 2K1C and 2K1C+IR groups. (g) Representative images of periodic acid-Schiff (PAS), MT and von Kossa staining of the kidney sections. Scale bars, 50 μm for PAS and von Kossa staining; 100 μm for MT staining. Kossa, von Kossa staining; TL, tibial length. Arrowheads indicate calcifications. *P<0.05 vs. the Control group; †P<0.05 vs. the 2K1C group. A full color version of this figure is available at the Hypertension Research journal online.
Impact of IR on renal structure in renovascular hypertensive rats
Stenosis of the unilateral renal artery develops into progressive atrophy of the stenotic kidney and compensatory hypertrophy of the contralateral kidney.14 Thus, we assessed the renal structure and function in the experimental groups. As shown in Figure 1f, the clipped kidney developed into renal atrophy in both the 2K1C and 2K1C+IR groups at 8 weeks after clipping. The clipped kidney weight to tibia length ratio was decreased in the 2K1C and 2K1C+IR groups, and the extent was not different between the 2K1C and 2K1C+IR groups (Figure 1f). The renal histology of the clipped kidney showed glomerulosclerosis, interstitial fibrosis and tubular atrophy in both the 2K1C and 2K1C+IR groups (Figure 1g). von Kossa staining of the clipped kidney showed diffuse calcifications in both the 2K1C and 2K1C+IR groups (Figure 1g).
However, unclipped kidney showed renal hypertrophy in the 2K1C and 2K1C+IR groups at 8 weeks after clipping, and the extent was less in the 2K1C+IR group (Figure 2a). The renal histology of the unclipped kidney revealed that glomerulosclerosis and interstitial fibrosis were increased in the 2K1C group at 8 weeks after clipping compared with those in the Control group, whereas these changes were attenuated in the 2K1C+IR group (Figures 2b–d). In addition, TUNEL staining showed an increase in apoptosis in the glomeruli and tubules of the 2K1C group at 8 weeks after clipping, whereas these increases were suppressed in the 2K1C+IR group compared with the 2K1C group (Figures 2b and e). Consistent with these findings, gene expression of collagen type I and collagen type III was increased in the unclipped kidney of the 2K1C group, whereas IR attenuated this increment (Figure 2f). Gene expression of CD68 and MCP-1 in the unclipped kidney was not different among the groups (Figure 2g). Collectively, these data indicate that IR attenuated the progression of compensatory hypertrophy in the unclipped kidney of 2K1C rats.

Impact of iron restriction (IR) on renal structure of the unclipped kidney in two-kidney one-clip (2K1C) rats. (a) Representative gross morphology of the unclipped kidney and unclipped kidney weight/TL ratio in the Control, 2K1C and 2K1C+IR groups. (b) Representative images of periodic acid-Schiff (PAS), Masson’s trichrome (MT), terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) and 4',6-diamidino-2-phenylindole staining (DAPI) staining of the kidney sections. Scale bars, 50 μm for PAS staining; 100 μm for MT, TUNEL and DAPI staining. White arrows indicate TUNEL-positive cells. Quantitative analysis of (c) the glomerular lesion score, (d) tubular lesion score and (e) TUNEL-positive cells per glomeruli and tubule in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). Renal gene expression of (f) collagen I, c ollagen III, (g) CD68 and MCP-1 in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). *P<0.05 vs. the Control group; †P<0.05 vs. the 2K1C group. MCP-1, monocyte chemoattractant protein-1; TL, tibial length. A full color version of this figure is available at the Hypertension Research journal online.
Impact of IR on renal function in renovascular hypertensive rats
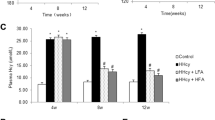
There was a trend toward an increase in proteinuria in the 2K1C group compared with the Control group at 7 weeks after clipping, whereas the increment was suppressed in the 2K1C+IR group compared with that in the 2K1C group (Figure 3a). Because oxidative stress is associated with the pathogenesis of RVHT,5, 6 we next assessed oxidative stress in the experimental groups. Dihydroethidium staining showed increased superoxide production in the unclipped kidney of the 2K1C group compared with that in the Control group, whereas IR attenuated this increment (Figure 3b). In addition, there was a trend toward an increase in urinary 8-OHdG and iron excretion in the 2K1C group compared with that in the Control group, whereas IR attenuated these increases in the 2K1C rats (Figures 3c and d). The tissue iron content in the unclipped kidney was decreased in the 2K1C+IR group compared with that in the other groups (Figure 3e).

Impact of iron restriction (IR) on renal function in two-kidney one-clip (2K1C) rats. (a) Urinary protein/creatinine excretion ratio in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). (b) Representative images of dihydroethidium (DHE) staining of the kidney sections and quantitative analysis of renal superoxide production in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). Scale bars, 100 μm. (c) Urinary 8-hydroxy-2'-deoxyguanosine (8-OHdG) excretion, (d) urinary iron excretion and (e) renal iron content in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). *P<0.05 vs. the Control group; †P<0.05 vs. the 2K1C group. A full color version of this figure is available at the Hypertension Research journal online.
Impact of IR on renal MR signaling in renovascular hypertensive rats
Because we have previously reported that IR attenuates renal MR signaling in 5/6 nephrectomized rats and normal Sprague–Dawley rats,11, 12 we next examined renal MR signaling in the unclipped kidney of the experimental groups. Importantly, there was a trend toward an increase in renal MR nuclear import in the 2K1C group compared with that in the Control group at 8 weeks after clipping, and it was decreased in the 2K1C+IR group compared with that in the 2K1C groups (Figure 4a). Similarly, there was a trend toward an increase in renal gene expression of SGK1, a key downstream effector of MR signaling, in the 2K1C group compared with that in the Control group, whereas the increase was suppressed in the 2K1C+IR group compared with that in the 2K1C group (Figure 4b). Urinary sodium excretion tended to be increased and urinary potassium excretion was decreased in the 2K1C+IR group compared with that in the other groups (Figures 4c and d). However, serum sodium and potassium levels and plasma aldosterone levels were not different among the groups (Table 1).

Impact of iron restriction (IR) on renal mineralocorticoid receptor (MR) signaling in two-kidney one-clip (2K1C) rats. (a) Renal nuclear expression of MR, (b) renal serum and glucocorticoid regulated kinase 1 (SGK1) gene expression, (c) urinary sodium excretion and (d) urinary potassium excretion in the Control, 2K1C and 2K1C+IR groups (n=6 in each group). *P<0.05 vs. the Control group; †P<0.05 vs. the 2K1C group. CREB, cAMP response element binding protein.
Impact of mild IR on hypertension and renal damage in renovascular hypertensive rats
As shown in Table 1, an iron-restricted diet induced severe anemia in 2K1C rats, and it may lead to beneficial effects on hypertension and renal damage in 2K1C rats. Thus, we further examined the effects of mild IR (30% iron-restricted diet) in 2K1C rats. The blood hemoglobin levels and body weight were slightly decreased in the 2K1C+mild IR group compared with those in the other groups at 8 weeks after clipping (Figures 5a and b). The increments of systolic blood pressure at 2 weeks after clipping were attenuated in the 2K1C+mild IR group compared with that in the 2K1C group (Figure 5c). In addition, proteinuria, glomerulosclerosis and renal interstitial fibrosis were suppressed in the 2K1C+mild IR group compared with that in the 2K1C group (Figures 5d–f).

Impact of mild iron restriction (IR) in two-kidney one-clip (2K1C) rats. (a) Blood hemoglobin levels, (b) body weight, (c) systolic blood pressure and (d) urinary protein/creatinine excretion ratio in the Control, 2K1C and 2K1C+mild IR groups (n=6 in each group). (e) Representative images of periodic acid-Schiff staining (PAS) and Masson’s trichrome (MT) staining of the kidney sections. Scale bars: 50 μm for PAS staining; 100 μm for MT staining. (f) Quantitative analysis of the glomerular lesion score and tubular lesion score in the Control, 2K1C and 2K1C+mild IR groups (n=6 in each group). *P<0.05 vs. the Control group; †P<0.05 vs. the 2K1C group. A full color version of this figure is available at the Hypertension Research journal online.
Discussion
To our knowledge, this study showed, for the first time, a novel effect of IR on hypertension and renal damage in RVHT. Hypertension is a major risk factor of cardiovascular death and is related to morbidity and mortality. Although essential hypertension is the most common form of hypertension, there is increasing recognition of secondary hypertension leading to morbidity and mortality in patients with hypertension. We have previously reported that IR prevents organ remodeling in several animal models of salt-sensitive hypertension.10, 11, 12 In this study, we wanted to assess the effect of IR on secondary hypertension induced by renal artery stenosis to evaluate whether dietary IR could attenuate the development of hypertension and organ remodeling in 2K1C rats.
Renal artery stenosis leads to both hypertension and chronic kidney disease.2 In this study, dietary IR attenuated the initial development of hypertension from the day of clipping in 2K1C rats. In addition, histological analyses showed that vascular remodeling, such as vascular hypertrophy and increased fibrosis, was suppressed in the 2K1C+IR group compared with that in the 2K1C group at 8 weeks after clipping. Because vascular remodeling is a chronic change, attenuation at the initial development of hypertension may lead to the progression of vascular remodeling. Furthermore, the heart rate and phosphorylation of eNOS at Ser1177 in the aorta were not different among the groups at 8 weeks after clipping. Oxidative stress is associated with the pathogenesis of RVHT.5, 6 In addition, MR signaling is associated with the progression of hypertension.15, 16 In this study, we found that IR attenuated superoxide production and MR nuclear import in the unclipped kidney, as well as urinary 8-OHdG excretion in 2K1C rats. Thus, IR may suppress the initial development of hypertension through the inhibition of oxidative stress and renal MR signaling.
By contrast, IR attenuated renal damage in 2K1C rats. This beneficial effect of IR on renal damage is consistent with the results previously reported in different models of renal disease.11, 12, 17, 18 Stenosis of the unilateral renal artery develops into progressive atrophy of the stenotic kidney and compensatory hypertrophy of the contralateral kidney, which might contribute to the pathogenesis of renovascular disease.14 In this study, IR did not affect renal atrophy in the clipped kidney, whereas compensatory hypertrophy of the contralateral kidney was attenuated in the 2K1C+IR group compared with that in the 2K1C group. The signaling pathways associated with fibrosis and inflammation are reported to be activated in the contralateral kidney following induction of renal artery stenosis.19 In the present study, IR attenuated gene expression of collagen type I and collagen type III but not that of CD68 and MCP-1 in the contralateral kidney. Therefore, IR may suppress compensatory hypertrophy of the contralateral kidney by the inhibition of fibrosis but not inflammation.
Iron is a vital element that is necessary for life. In the current study, we started the experiments using immature rats (5-week-old), and we observed lower body weight in the 2K1C+IR group compared with that in the other groups at 8 weeks after clipping. Meanwhile, mild IR (30% iron-restricted diet) slightly decreased body weight in 2K1C rats. Thus, these results indicate that iron is necessary for growth. By contrast, iron is a catalyst in the formation of reactive oxygen species, which induces cell and tissue damage. In fact, in both humans and experimental animal models of RVHT, increased oxidative stress is associated with the pathogenesis of RVHT.5, 6 In this study, we showed decreased superoxide production in the contralateral kidney and urinary 8-OHdG excretion in the 2K1C+IR group compared with that in the 2K1C group. Therefore, IR might attenuate the development of hypertension and renal damage in 2K1C rats through the inhibition of oxidative stress.
MR signaling contributes to the progression of hypertension and renal disease.15, 16 We have previously reported that IR attenuates renal MR signaling in normal Sprague–Dawley rats and 5/6 nephrectomized rats.11, 12 Similar to our previous results, we found that IR attenuated renal MR nuclear import and SGK1 gene expression in the unclipped kidney of 2K1C rats. These data suggest that IR attenuated the development of hypertension and renal damage through the inhibition of renal MR signaling, along with the inhibition of oxidative stress. However, oxidative stress is associated with renal MR activation.20 Thus, IR may suppress renal MR signaling through the attenuation of oxidative stress in 2K1C rats.
A recent clinical study has shown that higher blood hematocrit levels are associated with the incidence of hypertension in men.21 Meanwhile, long-term IR induces iron deficiency anemia. Actually, in this study, an iron-restricted diet induced severe anemia in 2K1C rats, and it may lead to the beneficial effects on hypertension and renal damage in 2K1C rats. Therefore, we further examined the effects of mild IR (30% iron-restricted diet) in this rat model. Mild IR slightly reduced blood hemoglobin levels; however, it attenuated the development of hypertension and renal damage in 2K1C rats, suggesting that IR could contribute to the preventive effects of hypertension and renal damage in 2K1C rats. Although severe anemia must be corrected, systemic iron balance should be considered for the pathogenesis of RVHT.
In conclusion, we demonstrated a novel effect of IR on hypertension and renal damage in 2K1C rats.
References
Derkx FH, Schalekamp MA . Renal artery stenosis and hypertension. Lancet 1994; 344: 237–239.
Goldblatt H, Lynch J, Hanzal RF, Summerville WW . Studies on experimental hypertension: I. the production of persistent elevation of systolic blood pressure by means of renal ischemia. J Exp Med 1934; 59: 347–379.
ASTRAL Investigators, Wheatley K, Ives N, Gray R, Kalra PA, Moss JG, Baigent C, Carr S, Chalmers N, Eadington D, Hamilton G, Lipkin G, Nicholson A, Scoble J . Revascularization versus medical therapy for renal-artery stenosis. N Engl J Med 2009; 361: 1953–1962.
Cooper CJ, Murphy TP, Cutlip DE, Jamerson K, Henrich W, Reid DM, Cohen DJ, Matsumoto AH, Steffes M, Jaff MR, Prince MR, Lewis EF, Tuttle KR, Shapiro JI, Rundback JH, Massaro JM, D'Agostino RB Sr, Dworkin LD, CORAL Investigators. Stenting and medical therapy for atherosclerotic renal-artery stenosis. N Engl J Med 2014; 370: 13–22.
Higashi Y, Sasaki S, Nakagawa K, Matsuura H, Oshima T, Chayama K . Endothelial function and oxidative stress in renovascular hypertension. N Engl J Med 2002; 346: 1954–1962.
Lerman LO, Nath KA, Rodriguez-Porcel M, Krier JD, Schwartz RS, Napoli C, Romero JC . Increased oxidative stress in experimental renovascular hypertension. Hypertension 2001; 37: 541–546.
Thurston H, Bing RF, Swales JD . Reversal of two-kidney one clip renovascular hypertension in the rat. Hypertension 1980; 2: 256–265.
al-Qattan KK, Johns EJ . A comparison of the actions of cilazapril in normal, dietary sodium-depleted and two-kidney, one clip Goldblatt hypertensive anaesthetised rats. J Hypertens 1992; 10: 423–429.
Tiradentes RV, Santuzzi CH, Claudio ER, Mengal V, Silva NF, Neto HA, Bissoli NS, Abreu GR, Gouvea SA . Combined Aliskiren and L-arginine treatment reverses renovascular hypertension in an animal model. Hypertens Res 2015; 38: 471–477.
Naito Y, Hirotani S, Sawada H, Akahori H, Tsujino T, Masuyama T . Dietary iron restriction prevents hypertensive cardiovascular remodeling in Dahl salt-sensitive rats. Hypertension 2011; 57: 497–504.
Naito Y, Fujii A, Sawada H, Hirotani S, Iwasaku T, Okuhara Y, Eguchi A, Ohyanagi M, Tsujino T, Masuyama T . Dietary iron restriction prevents further deterioration of renal damage in a chronic kidney disease rat model. J Hypertens 2013; 31: 1203–1213.
Naito Y, Fujii A, Sawada H, Hirotani S, Iwasaku T, Eguchi A, Ohyanagi M, Tsujino T, Masuyama T . Effect of iron restriction on renal damage and mineralocorticoid receptor signaling in a rat model of chronic kidney disease. J Hypertens 2012; 30: 2192–2201.
Naito Y, Fujii A, Sawada H, Oboshi M, Iwasaku T, Okuhara Y, Morisawa D, Eguchi A, Hirotani S, Masuyama T . Association between renal iron accumulation and renal interstitial fibrosis in a rat model of chronic kidney disease. Hypertens Res 2015; 38: 463–470.
Gobé GC, Axelsen RA, Searle JW . Cellular events in experimental unilateral ischemic renal atrophy and in regeneration after contralateral nephrectomy. Lab Invest 1990; 63: 770–779.
Pitt B . Mineralocorticoid receptor antagonists for the treatment of hypertension and the metabolic syndrome. Hypertension 2015; 65: 41–42.
Le Menuet D, Isnard R, Bichara M, Viengchareun S, Muffat-Joly M, Walker F, Zennaro MC, Lombès M . Alteration of cardiac and renal functions in transgenic mice overexpressing human mineralocorticoid receptor. J Biol Chem 2001; 276: 38911–38920.
Naito Y, Sawada H, Oboshi M, Fujii A, Hirotani S, Iwasaku T, Okuhara Y, Eguchi A, Morisawa D, Ohyanagi M, Tsujino T, Masuyama T . Increased renal iron accumulation in hypertensive nephropathy of salt-loaded hypertensive rats. PLoS ONE 2013; 8: e75906.
Sawada H, Naito Y, Oboshi M, Iwasaku T, Okuhara Y, Morisawa D, Eguchi A, Hirotani S, Masuyama T . Iron restriction inhibits renal injury in aldosterone/salt-induced hypertensive mice. Hypertens Res 2015; 38: 317–322.
Cheng J, Zhou W, Warner GM, Knudsen BE, Garovic VD, Gray CE, Lerman LO, Platt JL, Romero JC, Textor SC, Nath KA, Grande JP . Temporal analysis of signaling pathways activated in a murine model of two-kidney, one-clip hypertension. Am J Physiol Renal Physiol 2009; 297: F1055–F1068.
Nagase M, Matsui H, Shibata S, Gotoda T, Fujita T . Salt-induced nephropathy in obese spontaneously hypertensive rats via paradoxical activation of the mineralocorticoid receptor: role of oxidative stress. Hypertension 2007; 50: 877–883.
Jae SY, Kurl S, Laukkanen JA, Heffernan KS, Choo J, Choi YH, Park JB . Higher blood hematocrit predicts hypertension in men. J Hypertens 2014; 32: 245–250.
Acknowledgements
We gratefully acknowledge the technical assistance of Sachi Ito and Kyoko Ogata for her excellent support. This study was supported by a Grant-in-Aid for Scientific Research (C) JSPS KAKENHI Grant No. 25460919 and No. 16K09273 and grants from The Salt Science Research Foundation (No. 1544) and Takeda Science Foundation (to YN). This study was partly supported by a Grant-in-Aid for Scientific Research (C) (JSPS KAKENHI Grant No. 15K09148) (to TT).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Oboshi, M., Naito, Y., Sawada, H. et al. Attenuation of hypertension and renal damage in renovascular hypertensive rats by iron restriction. Hypertens Res 39, 832–839 (2016). https://doi.org/10.1038/hr.2016.93
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hr.2016.93
Keywords
This article is cited by
-
Ferritin modifies the relationship between inflammation and arterial stiffness in hypertensive patients with different glucose tolerance
Cardiovascular Diabetology (2020)
-
Reply to ‘Iron restriction in renovascular hypertension’
Hypertension Research (2017)
-
Iron restriction in renovascular hypertension
Hypertension Research (2017)
-
Altered expression of intestinal duodenal cytochrome b and divalent metal transporter 1 might be associated with cardio-renal anemia syndrome
Heart and Vessels (2017)
-
Novel insights into renovascular hypertension and cardio-renal protection by iron restriction
Hypertension Research (2016)
