
Abstract
The colonization of Eurasia by early humans is a key event after their spread out of Africa, but the nature, timing and ecological context of the earliest human occupation of northwest Europe is uncertain and has been the subject of intense debate1. The southern Caucasus was occupied about 1.8 million years (Myr) ago2, whereas human remains from Atapuerca-TD6, Spain (more than 780 kyr ago)3 and Ceprano, Italy (about 800 kyr ago)4 show that early Homo had dispersed to the Mediterranean hinterland before the Brunhes–Matuyama magnetic polarity reversal (780 kyr ago). Until now, the earliest uncontested artefacts from northern Europe were much younger, suggesting that humans were unable to colonize northern latitudes until about 500 kyr ago5,6. Here we report flint artefacts from the Cromer Forest-bed Formation at Pakefield (52° N), Suffolk, UK, from an interglacial sequence yielding a diverse range of plant and animal fossils. Event and lithostratigraphy, palaeomagnetism, amino acid geochronology and biostratigraphy indicate that the artefacts date to the early part of the Brunhes Chron (about 700 kyr ago) and thus represent the earliest unequivocal evidence for human presence north of the Alps.
Similar content being viewed by others

Main
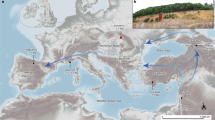
The Cromer Forest-bed Formation (CF-bF), exposed discontinuously for a distance of more than 80 km along the North Sea coast of eastern England, has long been famous for its early Middle Pleistocene fossils7,8,9,10,11. Spectacular finds include many extinct large mammals, molluscs, beetles, remains of fruits, seeds and even trees from which the deposits get their name. Recent work on vertebrate and molluscan faunas has shown that the CF-bF is much more complex than previously realized and includes evidence for at least six distinct temperate phases between about 780 and 450 kyr ago12. The CF-bF sediments are primarily organic detritus muds and sands laid down within channels and on the floodplains of rivers, which drained central and eastern England before ice sheets invaded the area about 450 kyr ago13. During this time, Britain was connected to what is now mainland Europe, East Anglia being located at the southwestern margin of a large coastal embayment around the subsiding North Sea basin. The flint artefacts found at Pakefield (52° 25.9′ N, 1° 43.8′ W) were recovered from river sediments, with a significant quartz and quartzite component, which formed the floodplain of the lower reaches of the erstwhile Bytham River that drained the English Midlands at this time (Fig. 1)13,14.

All known Anglian/Elsterian and earlier archaeological sites are shown, and their concentration in southern England is highlighted. These sites can now be shown to provide a record of intermittent early human occupation of about 300 kyr duration. Human remains are known from Mauer and Boxgrove. M, Miesenheim I; W, Westbury-sub-Mendip; WR, West Runton; WW, Waverley Wood. For clarity, the names of rivers or geographical features are italicized and site names are in Roman font.
Despite two centuries of investigation, no convincing artefacts have hitherto been found stratified within the CF-bF. However, several excavations in the recently re-exposed coastal sections of CF-bF (and associated deposits) between Pakefield and Kessingland11,14,15,16 resulted in the discovery of 32 worked flints (length more than 20 mm), including a simple flaked core, a crudely retouched flake and a quantity of waste flakes (Fig. 2). These artefacts come from clear stratigraphical contexts and are associated with a wealth of evidence that allows a rare opportunity to reconstruct the environment inhabited by the humans who made the tools (see Supplementary Table 1). The artefacts, all in very sharp condition, are made of good quality black flint. Unworked surfaces, where present, are water-worn, indicating that the raw material might have been collected from the adjacent river channel. The assemblage lacks formal tools and is thus consistent with a Mode 1 technology (that is, flakes, pebble tools and choppers made with hard hammers17), but this interpretation must remain provisional in view of the small sample size.

a, Core, partly alternate hard-hammer flaking, with several incipient cones of percussion on platforms. b, Retouched flake. c–j, Hard hammer-struck flakes, with previous removals on dorsal surface, often from the same direction. b, ‘Rootlet bed’ (PaB); c, laminated silts (PaCii); a, d–j, ‘Unio-bed’ (PaCi).
The artefacts came from four different contexts within an interglacial infill of a channel incised into Early Pleistocene marine sediments and overlain by a sequence of marine sands, glaciofluvial sediments and Lowestoft Till (Fig. 3). The oldest artefact was found in the upper levels of the estuarine silts (Fig. 3) containing marine and brackish-water ostracods, foraminifera and sparse marine mammals (including dolphin and walrus). All other artefacts were found in the CF-bF, which comprises the ‘Rootlet bed’, ‘Unio-bed’ and ‘laminated silts’14,15,16. Two artefacts were found in overbank sediments with well-developed soil features including numerous fossil root-casts and pedogenic carbonate nodules (‘Rootlet bed’). Most of the artefacts (n = 28) came from a lag gravel (‘Unio-bed’) at the base of the laminated silts that fill the channel cut into the overbank sediments, but a single flake (Fig. 2c) was found in the laminated silts at the edge of the channel. Sediments overlying the channel infill were originally interpreted as glaciofluvial deposits attributed to the North Sea Drift Formation (Marine Isotope Stage (MIS) 12; refs 11, 18; Fig. 3b), thus implying a long hiatus between the CF-bF and the glacial sequence. A new interpretation14 suggests that a more complete sequence is represented (Fig. 3b) by the complex succession of marine sands, Bytham River sands and gravels, and glaciofluvial sands and gravels from the Happisburgh Glaciation, which are separated from the CF-bF by a periglacial interval indicated by the presence of frost-cracks. Another marine deposit overlies these sands and gravels and the whole sequence is capped by the Lowestoft Till Member, deposited by the Anglian Glaciation11, when ice sheets reached their maximum extent in Britain.

a, Coastal sections between Pakefield and Kessingland. b, Geological profile and interpretation.
Fossils from sediments that contain the artefacts indicate that the local climate was significantly different from that of the present day in terms of temperature and seasonality of precipitation. Plant (Trapa natans, Salvinia natans and Corema album) and beetle (Cybister lateralimarginalis, Oxytelus opacus and Valgus hemipterus) remains include several thermophilous species no longer living in Britain, and the presence of Hippopotamus and frost-sensitive insects and plants implies warmer summers and mild winters. Mutual Climatic Range (MCR)19 analysis of the beetle assemblage enables quantitative estimates to be made of the thermal climate at that time. These suggest that the mean temperature of the warmest month (July) was between 18 and 23 °C and the mean temperature of the coldest months (January/February) was between -6 and +4 °C. Pedogenic carbonate nodules in the ‘Rootlet bed’ indicate an annual moisture deficit, whereas their isotopic composition reflects intense soil moisture evaporation during their formation20. The combination of warmer summer and winter temperatures together with a strongly seasonal precipitation regime is indicative of a warm, seasonally dry Mediterranean climate.
Both the insects and plants indicate the presence of marshy ground with extensive reedy vegetation and alder trees adjacent to a meandering river with shallow riffles and deeper pools. Oak woodland grew on drier ground with open grassland nearby. This mixture of habitats supported a variety of large browsing and grazing mammals dominated by Mammuthus trogontherii, Stephanorhinus hundsheimensis, Megaloceros savini, M. dawkinsi and Bison cf. schoetensacki, and their predators and scavengers (Homotherium sp., Panthera leo, Canis lupus (small) and Crocuta crocuta). The floodplain would have provided a resource-rich environment for early humans, with a range of plant and animal resources. An additional attraction, in an area where good quality flint would have been scarce, was the flint-rich river gravels, which provided the raw material for tool manufacture.
Pollen analysis indicates that the channel infill accumulated during an interglacial with regional vegetation dominated by broad-leaf woodland that included Carpinus (hornbeam) in its later part11. These sediments were originally correlated with those at the Cromerian stratotype at West Runton, Norfolk, 60 km to the northwest, on the basis of palynology11 and malacology (for example the presence of the extinct freshwater mollusc Valvata goldfussiana12). This suggestion is broadly supported by the extent of racemization and the pattern of amino acid decomposition in the intracrystalline fraction of Bithynia opercula (see Methods and Supplementary Information). However, although these sites are close in age, several lines of evidence indicate that they are not exactly contemporary. West Runton lacks the important component of southern thermophile plants and beetles seen at Pakefield. Moreover, several large mammals, such as Hippopotamus, Megaloceros dawkinsi and Palaeoloxodon antiquus (the latter recorded only from unstratified material) known from Pakefield/Kessingland have never been found at West Runton, despite the much more extensive collections from that site16. Furthermore, the occurrence at Pakefield of two vole species of the genus Mimomys, M. savini and M. aff. pusillus (see Supplementary Information), of which only the former is known from West Runton, is also consistent with a difference in age. M. pusillus is known from the Early Pleistocene to its latest occurrence in the early Middle Pleistocene Ilynian Complex (European Russia)21. As the latter is overlain by the Don Till (Donian), correlated with MIS 16 (ref. 21), this suggests a minimum age for the Pakefield ‘Unio-bed’.
This evidence indicates that sediments containing the artefacts belong to an interglacial in the early part of the ‘Cromerian Complex’10. This interpretation is supported by independent evidence derived from the lithostratigraphy. A maximum age is indicated by palaeomagnetic data from the laminated silts, which show normal polarity14, consistent with the early part of the Brunhes Chron. A minimum age is indicated by the overlying Lowestoft Till Member, conventionally thought to have been emplaced during MIS 12 (refs 13, 18). The sediments between the CF-bF and the till have been traditionally interpreted as being of glaciofluvial origin, deposited during the Anglian Stage11,17. A longer chronology has recently been proposed on the basis of sedimentological evidence suggesting that the CF-bF may be separated from the MIS 12 till by two separate high sea-stands and two cold episodes14 (Fig. 3b). By using the premise that temperate-climate marine deposits and cold-climate aggradations in the lower parts of large, temperate latitude rivers can be matched, respectively, to the peaks and troughs of orbitally tuned MIS cycles22, the archaeology can be dated to MIS 17 (about 680 kyr ago) at the very youngest. If evidence for ice-sheet extension across eastern England during MIS 16 is valid23, then an additional temperate/cold cycle is required and the archaeology at Pakefield could be as old as the later part of MIS 19 (about 750 kyr ago).
The oldest human fossils in northwest Europe are from Mauer6, Germany and Boxgrove24, UK, where they are part of vertebrate assemblages that include the water vole Arvicola terrestris cantiana. All other northwestern European sites containing early Middle Pleistocene archaeology in association with rich small-mammal assemblages (for example, Miesenheim I in Germany, and Westbury-sub-Mendip and Waverley Wood in the UK)6,17,25, have similarly yielded Arvicola (with unrooted molars) rather than Mimomys (its ancestor with rooted molars). These early Middle Pleistocene archaeological sites with Arvicola have all been correlated with MIS 13, the basis for the belief in a short chronology for human occupation in this region6. The discovery at Pakefield of unequivocal artefacts in beds yielding Mimomys demonstrates a much longer human occupation of northwest Europe, pre-dating other evidence by as much as 200 kyr. There has been much discussion about what additional social, technological or physiological adaptations humans would have required to colonize northwest Europe compared with their occupation farther south5,26. The Mediterranean climate reconstructed for the archaeological levels at Pakefield suggests that these pioneers were able to spread northwards in familiar climatic conditions, using their existing adaptations.
Methods
A new technique of amino acid analysis has been developed for geochronological purposes27 that combines a new reverse-phase–high-pressure liquid chromatography (RP–HPLC) method of analysis28 with the isolation of an intracrystalline fraction of amino acids by treatment with bleach29. This combination of techniques results in the analysis of d/l values of multiple amino acids from the chemically protected protein within the biomineral, enabling both smaller sample sizes and increased reliability of the analysis. The calcitic structure of the opercula of Bithynia (a genus of freshwater prosobranch gastropods) has been found to provide the most robust repository for the amino acids yet studied.
Amino acid racemization analyses were undertaken with these procedures on three individual samples of Bithynia troscheli opercula (NEaar 1711–1712, 1968) from the laminated silts (PaCii) and four samples of B. troscheli opercula (NEaar 2146–2167) from the Unio-bed (PaCi). Each operculum was powdered and bleached for 48 h with 12% NaOCl. Two subsamples were taken: one fraction was directly demineralized and the free amino acids were analysed (referred to as the ‘free’ fraction, F), and the second was treated with 7 M HCl under N2 at 110 °C for 24 h (referred to as the ‘hydrolysed’ fraction, H*) to release the peptide-bound amino acids, thus yielding the ‘total’ amino acid concentration. Samples were then dried with a centrifugal evaporator and rehydrated for RP–HPLC analysis with 0.01 mM l-homo-arginine as an internal standard.
The amino acid compositions were analysed in duplicate by RP–HPLC using fluorescence detection, following the method of ref. 28. A 2 µl sample was injected and mixed online with 2.2 µl of derivitizing reagent (260 mM n-iso-l-butyryl l-cysteine (IBLC) and 170 mM o-phthaldialdehyde (OPA) in 1 M potassium borate buffer, adjusted to pH 10.4 with potassium hydroxide pellets). The amino acids were separated on a C18 HyperSil BDS column (5 mm × 250 mm) at 25 °C with a gradient elution of three solvents: sodium acetate buffer (solvent A; 23 mM sodium acetate trihydrate, 1.5 mM sodium azide, 1.3 µM EDTA, adjusted to pH 6.00 ± 0.01 with 10% acetic acid and sodium hydroxide), methanol (solvent C) and acetonitrile (solvent D). The l and d isomers of ten amino acids were routinely analysed. During preparative hydrolysis both asparagine and glutamine undergo rapid irreversible deamination to aspartic acid and glutamic acid, respectively29. It is therefore not possible to distinguish between the acidic amino acids and their derivatives; they are therefore reported together as Asx and Glx.
On the basis of protein decomposition (racemization and degradation of amino acids) we conclude that the results are consistent with attribution to an early part of the ‘Cromerian Complex’27. All opercula from Pakefield are significantly older (95% confidence) than those analysed from Waverley Wood, but they cannot be statistically separated from opercula from the type Cromerian West Runton Freshwater Bed. One sample from the Unio-bed (NEaar 2166) seems to be slightly younger than the others. The amino acid data therefore indicate that the Pakefield opercula are at least as old as those from West Runton (see Supplementary Table 2).
References
Dennell, R. Dispersal and colonisation, long and short chronologies: how continuous is the Early Pleistocene record for hominids outside Africa? J. Hum. Evol. 45, 421–440 (2003)
Gabunia, L. et al. Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: Taxonomy, geological setting, and age. Science 288, 1019–1025 (2000)
Carbonell, E. et al. Lower Pleistocene hominids and artifacts from Atapuerca-TD6 (Spain). Science 269, 826–829 (1995)
Manzi, G. Human evolution at the Matuyama–Brunhes boundary. Evol. Anthropol. 13, 11–24 (2004)
Gamble, C. The Palaeolithic Societies of Europe (Cambridge Univ. Press, Cambridge, 1999)
Dennell, R. & Roebroeks, W. The earliest colonization of Europe: the short chronology revisited. Antiquity 70, 535–542 (1996)
Newton, E. T. The Vertebrata of the Forest Bed Series of Norfolk and Suffolk (Memoirs of the Geological Survey, United Kingdom, London, 1882)
Reid, C. The Geology of the Country around Cromer (Memoirs of the Geological Survey, England and Wales, London, 1882)
Stuart, A. J. in The early Middle Pleistocene in Europe (ed. Turner, C.) 9–24 (Balkema, Rotterdam, 1996)
Turner, C. in The early Middle Pleistocene in Europe (ed. Turner, C.) 295–317 (Balkema, Rotterdam, 1996)
West, R. G. The pre-glacial Pleistocene of the Norfolk and Suffolk Coasts (Cambridge Univ. Press, Cambridge, 1980)
Preece, R. C. Molluscan evidence for differentiation of interglacials within the ‘Cromerian Complex’. Quat. Sci. Rev. 20, 1643–1656 (2001)
Rose, J., Moorlock, B. S. P. & Hamblin, R. J. O. Pre-Anglian fluvial and coastal deposits in Eastern England: lithostratigraphy and palaeoenvironments. Quat. Int. 79, 5–22 (2001)
Lee, J. R., Rose, J., Candy, I. & Barendregt, R. Sea-level changes, river activity, soil development and glaciation around the western margins of the southern North Sea basin during the Early and early Middle Pleistocene: evidence from Pakefield, Suffolk, UK. J. Quat. Sci. (in the press)
Blake, J. H. The Geology of the Country near Yarmouth and Lowestoft. (Memoirs of the Geological Survey, England and Wales, London, 1890, relating to Horizontal Section 128).
Stuart, A. J. & Lister, A. M. The mammalian faunas of Pakefield/Kessingland and Corton, Suffolk, U.K.: evidence for a new temperate episode in the British early Middle Pleistocene. Quat. Sci. Rev. 20, 1677–1692 (2001)
Wymer, J. The Lower Palaeolithic Occupation of Britain (Wessex Archaeology & English Heritage, Salisbury, 1999)
Bowen, D. Q. (ed.) A New Correlation of Quaternary Deposits in the British Isles (Geol. Soc. Spec. Rep. 23, London, 1999).
Atkinson, T. C., Briffa, K. A. & Coope, G. R. Seasonal temperatures in Britain during the past 22,000 years, reconstructed using beetle remains. Nature 325, 587–592 (1987)
Candy, I., Rose, J. & Lee, J. R. A seasonally ‘dry’ interglacial climate in eastern England during the early Middle Pleistocene: palaeopedological and stable isotopic evidence from Pakefield, UK. Boreas (in the press)
Pevzner, M., Vangengeim, E. & Tesakov, A. Quaternary subdivisions of Eastern Europe based on vole evolution. Boll. Soc. Paleont. Ital. 40, 269–274 (2001)
Bassinot, F. C. et al. The astronomical theory of climate and the age of the Brunhes–Matuyama magnetic reversal. Earth Planet. Sci. Lett. 126, 91–108 (1994)
Lee, J. R., Rose, J., Hamblin, R. J. O. & Moorlock, B. S. P. Dating the earliest lowland glaciation of eastern England: a pre-MIS 12 early Middle Pleistocene glaciation. Quat. Sci. Rev. 23, 1551–1566 (2004)
Roberts, M. B. & Parfitt, S. A. Boxgrove: A Middle Pleistocene Hominid Site at Eartham Quarry, Boxgrove, West Sussex. (English Heritage, London, 1999)
Bishop, M. J. Earliest record of Man's presence in Britain. Nature 253, 95–97 (1975)
Roebroeks, W. Hominid behaviour and the earliest occupation of Europe: an exploration. J. Hum. Evol. 41, 437–461 (2001)
Penkman, K. E. H. Amino Acid Geochronology: A Closed System Approach to Test and Refine the UK Model. Thesis, Univ. Newcastle (2005)
Kaufman, D. S. & Manley, W. F. A new procedure for determining DL amino acid ratios in fossils using reverse phase liquid chromatography. Quat. Sci. Rev. 17, 987–1000 (1998)
Sykes, G. A., Collins, M. J. & Walton, D. I. The significance of a geochemically isolated intracrystalline organic fraction within biominerals. Org. Geochem. 23, 1039–1065 (1995)
Gibbard, P. L. The history of the great northwest European rivers during the past three million years. Phil. Trans. R. Soc. Lond. B 318, 559–602 (1988)
Acknowledgements
We thank N. M. Ashton, A. P. Currant, P. L. Gibbard, N. R. Larkin, M. D. Lewis, S. G. Lewis, W. Roebroeks, J. R. Stewart, C. Turner and A. Charlton for help in the field and/or discussions; T. Lawson, who found the first putative artefact in 2000; and P. Crabb (NHM photo unit) for photographs of the vole molars. A. W. Gentry commented on the bovid remains. R.W.B. acknowledges support from NSERC (Canada) and laboratory assistance provided by R. Enkin and J. Baker at the Geological Survey of Canada-Pacific. J.R.L. publishes with the permission of the Executive Director, British Geological Survey (NERC). We thank the Leverhulme Trust, which supported this work with grants to the Ancient Human Occupation of Britain (AHOB) project (http://www.nhm.ac.uk/hosted_sites/ahob/), and the project ‘Mammoth, Vertebrate Fauna and Palaeoenvironment of the Cromerian Stratotype’ (A.M.L. and A.J.S.). Amino acid geochronology was funded by English Heritage (Aggregates Levy Sustainability Fund) and NERC. This work was undertaken while J.R.L. was in receipt of a NERC studentship at Royal Holloway, University of London, and the British Geological Survey. Author Contributions S.A.P., J.R. and A.J.S. coordinated research on the site, and S.A.P. wrote the article, with major contributions from R.C.P., A.J.S., J.R. and C.B.S. The stratigraphical and lithological sequence was described by J.R.L. and J.R., and I.C. investigated the sediment geochemistry. R.M. and P.D. recovered most of the artefacts and mammalian fauna; the artefacts were figured and described by J.J.W. The large mammals were studied by M.B., A.J.S. and A.M.L., and S.A.P. analysed the small mammal fauna. K.E.H.P. and M.J.C. are responsible for the amino acid geochronology and R.W.B. undertook the palaeomagnetic investigations. Beetles were analysed by G.R.C., molluscs by R.C.P., plant macrofossils by M.H.F., and ostracods and foraminifera by J.E.W. R.S. made major contributions in the field and in drawing the figures.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Reprints and permissions information is available at npg.nature.com/reprintsandpermissions. The authors declare no competing financial interests.
Supplementary information
Supplementary Table 1 (XLS 107 KB)
Biological remains from Pleistocene deposits at Pakefield and Kessingland, Suffolk, UK.
Supplementary Table 2 (DOC 45 KB)
Amino acid composition of Bithynia troscheli opercula from Pakefield (laminated silts, lithofacies PaCi and ‘Unio-bed’, PaCii), Waverley Wood and the type Cromerian, West Runton Freshwater Bed.

Supplementary Figure 1 (JPG 435 KB)
First lower molar of Mimomys aff. pusillus compared with that of M. savini (Pakefield ‘Unio-bed’). (a, c, e) (BMNH. M 63567, ex R. Mutch collection 1108.1, ‘Unio-bed’) compared with that of M. savini (b, d, f) (BMNH M. 63568, ex R. Mutch collection 1242.2, ‘Unio-bed’). a, b) occlusal view; c, d) lingual view; e, f) labial view. Scale bar = 1 mm.
Science Media Centre Briefing: Streaming flash videos - Earliest Record of Human Activity in Northern Europe
Early humans were living in Britain as much as 700,000 years ago, according to research in this week’s Nature. A newly discovered collection of stone tools in East Anglia has shown that human activity was present in Northern Europe 200,000 years earlier than scientists thought. This work challenges the current understanding of how humans moved from Southern Europe to colonise areas of Northern Europe – including the UK.
Background Information: Early humans were living in Britain as much as 700,000 years ago, according to research in this week’s Nature. A newly discovered collection of stone tools in East Anglia has shown that human activity was present in Northern Europe 200,000 years earlier than scientists thought. This work challenges the current understanding of how humans moved from Southern Europe to colonise areas of Northern Europe – including the UK.
You will need the Flash browser plugin to view this coverage. Download the Flash Plugin here: https://www.macromedia.com/go/getflashplayer
Rights and permissions
About this article
Cite this article
Parfitt, S., Barendregt, R., Breda, M. et al. The earliest record of human activity in northern Europe. Nature 438, 1008–1012 (2005). https://doi.org/10.1038/nature04227
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature04227
This article is cited by
-
Late Early to late Middle Pleistocene medium-sized deer from the Italian Peninsula: implications for taxonomy and biochronology
Palaeobiodiversity and Palaeoenvironments (2024)
-
Brief interviews with hideous stone: a glimpse into the butchery site of Isernia La Pineta — a combined technological and use-wear approach on the lithic tools to evaluate the function of a Lower Palaeolithic context
Archaeological and Anthropological Sciences (2023)
-
The Technological Behaviours of Homo antecessor: Core Management and Reduction Intensity at Gran Dolina-TD6.2 (Atapuerca, Spain)
Journal of Archaeological Method and Theory (2023)
-
Investigating variability in the frequency of fire use in the archaeological record of Late Pleistocene Europe
Archaeological and Anthropological Sciences (2022)
-
Lower and Early Middle Palaeolithic of Southern Britain: the Evidence from the River Test
Journal of Paleolithic Archaeology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}