
Abstract
Previously known only from isolated teeth and lower jaw fragments recovered from the Cretaceous and Palaeogene of the Southern Hemisphere, the Gondwanatheria constitute the most poorly known of all major mammaliaform radiations. Here we report the discovery of the first skull material of a gondwanatherian, a complete and well-preserved cranium from Upper Cretaceous strata in Madagascar that we assign to a new genus and species. Phylogenetic analysis strongly supports its placement within Gondwanatheria, which are recognized as monophyletic and closely related to multituberculates, an evolutionarily successful clade of Mesozoic mammals known almost exclusively from the Northern Hemisphere. The new taxon is the largest known mammaliaform from the Mesozoic of Gondwana. Its craniofacial anatomy reveals that it was herbivorous, large-eyed and agile, with well-developed high-frequency hearing and a keen sense of smell. The cranium exhibits a mosaic of primitive and derived features, the disparity of which is extreme and probably reflective of a long evolutionary history in geographic isolation.
Similar content being viewed by others

Main
Mammalia Linnaeus, 1758
Allotheria Marsh, 1880
Gondwanatheria Mones, 1987
Sudamericidae Scillato-Yané and Pascual, 1984
Vintana sertichi gen. et sp. nov.
Etymology. Vintana (Malagasy), luck, in reference to the circumstances of discovery of the holotype specimen. Species name after Joseph Sertich, discoverer of UA 9972.
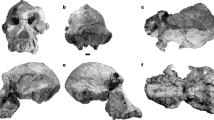
Holotype and only known specimen. A complete and well-preserved cranium, University of Antananarivo (UA) 9972 (Fig. 1; Supplementary Videos 1, 2, 3).

a–e, Holotypic specimen, UA 9972, in right lateral (a), dorsal (b), ventral (c), anterior (d) and posterior (e) views, with micro-computed tomography-based, digitally-rendered image on the left and line drawing reconstruction on the right in each pair. Hypothetical incisor crowns shown in part a reconstruction only, and the last three upper molariforms (MF) on both sides and alveoli for the single premolariform tooth (PMF) and MF1 in part c reconstruction only.
Locality and horizon. Locality MAD10-24, Upper Cretaceous (Maastrichtian; 72.1–66.0 Myr ago) Lac Kinkony Member, Maevarano Formation, Mahajanga Basin, northwestern Madagascar1.
Diagnosis. Taxon differs from all other gondwanatherians in its large size and in exhibiting wear features on molariform tooth crowns indicating a distobuccal (rather than strictly distal) power stroke of the chewing cycle. Full diagnosis in Supplementary Information.
Gondwanatherians are an enigmatic mammalian clade previously represented by only seven valid monotypic genera from the Cretaceous and Palaeogene of South America, Africa, India, Madagascar and the Antarctic Peninsula2,3. With the exception of a few dentary fragments, gondwanatherians were previously known only from isolated teeth3. No cranial or postcranial material has been assigned to the Gondwanatheria until now, a severe limitation that has left their phylogenetic position within Mammaliaformes uncertain and controversial. Virtually nothing is known of their life habits, aside from inferences that at least the larger sudamericids were herbivorous and ingested an abrasive diet4,5,6 and that ferugliotheriids were omnivorous5. The cranium described here, from the Late Cretaceous of Madagascar, is remarkably complete and well preserved and only the third known occurrence of a mammaliaform cranium from the Cretaceous of Gondwana7,8. It provides an unprecedented opportunity to more reliably assess gondwanatherian relationships and to analyse various aspects of gondwanatherian palaeobiology.
Dental features
UA 9972 represents the first instance in which upper teeth of a gondwanatherian mammal are associated in gnathic material. These teeth provide the foundation for assigning the specimen to a new genus and species and the opportunity to more comprehensively evaluate dental function and diet in gondwanatherians.
Vintana sertichi has an upper dentition consisting of two incisors, no canine, one premolariform tooth, and four molariform teeth in each quadrant. Although the incisors themselves are not preserved, there are two long, curved alveoli in each premaxilla for enlarged, laterally compressed, procumbent, and probably ever-growing incisors that were well separated from the cheek teeth by a long diastema (Figs 1a, c and 2a, b). Based on its alveoli, the single, two-rooted premolariform tooth appears to have been small; neither the left nor right crowns are preserved in UA 9972 (Figs 1c and 2c). Of the eight upper molariform teeth (MF) present in life in V. sertichi (four on each side), four heavily worn representatives (left MF2–4, right MF3) are preserved in UA 9972 (Fig. 2c). The molariform cheek teeth have several salient characteristics: large size, hypsodont crowns (extremely worn in UA 9972), quadrangular occlusal profiles, occlusal surfaces worn essentially flat (with heaviest wear in more mesial molariforms, indicating a mesial-to-distal eruption sequence), numerous cementum-filled infundibula, cementum-filled furrows that invaginate from the buccal side but do not extend to the base of the crown, and multiple short roots supporting the periphery of the base of each crown (Fig. 2c, d; Supplementary Videos 4 and 5). The molariform teeth are also exceptional in that their occlusal surfaces face laterally as much as they do ventrally (Figs 1c, d and 2c).

a, b, Lateral (a) and dorsal (b) views of snout region developed from micro-computed tomography data showing position, size and orientation of alveoli of the mesial (red) and distal (blue) incisors. Dashed lines in a indicate hypothetical outlines of right incisor crowns. c, Ventral (occlusal) view of upper cheek-tooth dentition showing preserved right third molariform (MF3) and left second, third and fourth molariforms (MF2–4) and positions of alveoli for other cheek teeth, including the single right premolariform (PMF). d, Occlusal view of reconstructed left MF2–4 showing distribution of cementum-filled islets, furrows and synclines.
The enamel microstructure of the molariform teeth of V. sertichi retains many of the plesiomorphic characteristics of mammaliaform prismatic enamel (for example, single-layered schmelzmuster; non-decussating, small prisms). However, V. sertichi appears to be derived in possessing modified radial enamel consisting of prisms separated by prominent interrow sheets of interprismatic matrix (Extended Data Fig. 1), thus resembling other gondwanatherians from the Late Cretaceous of Madagascar9,10 and India9.
Craniofacial features
The well-preserved and complete nature of UA 9972 permits the first insight into the craniofacial morphology of a gondwanatherian mammal. Superficially striking are its short, highly vaulted cranium, large orbits, elongated jugal flanges on widely flaring zygomatic arches, and strong klinorhynchy (Fig. 1; Supplementary Videos 1, 2, 3). However, more detailed examination reveals an array of primitive features reminiscent of the most basal mammaliaforms, or even non-mammaliaform cynodonts, coupled with highly derived features unknown in any other Mesozoic mammaliaform.
The snout of Vintana exhibits a number of features that are unique among mammaliaforms (Fig. 1; Extended Data Figs 2a, b; 3a), including the retention of a septomaxilla with both a large posterodorsal facial process and an intranarial process. The lacrimal bone is enormous and extends anteriorly to contact both the septomaxilla and premaxilla. The lacrimal and palatine bones of Vintana contribute significantly to the nasal cavity, thereby demonstrating in striking fashion that the identity of adult bones comprising the walls of the cavity varies considerably among mammaliaform taxa. Micro-computed tomography allows the confident identification of a range of nasal cavity structures that have been difficult or impossible to visualize in other Mesozoic mammaliaforms. These include remnants of two turbinal elements, the crista semicircularis and the first ethmoturbinal (including both its lateral and vertical roots), which mark the anterior and posterior boundaries of the lateral recess (fetal pars intermedia). Also clearly identifiable are the basal lamellae of the nasoturbinal and those from which the remaining ethmoturbinals emanated (on the frontal and palatine bones), the peripheral edges of the cribriform plate (on the frontal bone, separating the nasal cavity from the braincase), the posterior transverse lamina (on the palatine and orbitosphenoid bones, separating the cupular recess from the nasopharyngeal canal), and the nasolacrimal canal/groove. Clear evidence of a maxilloturbinal is not preserved. The inferred presence of well-developed turbinal structures in Vintana bears witness to the extraordinary conservatism of mammaliaform internal nasal anatomy (despite its unusual bony composition) but is also consistent with the relatively massive size of the olfactory bulbs and the deep, long (∼60% of cranial length) nasal cavity.
No Mesozoic mammaliaform has a jugal as enlarged, and with as massive a ventral flange, as that of Vintana (Fig. 1). The palate of Vintana is very narrow, has a rugose texture, and lacks vacuities or any sizable foramina (other than the incisive foramina anteriorly). Furthermore, the palatines are fused in the midline and extend forward to contact the premaxillae, thus excluding the maxillae from contacting one another in the midline; this feature is unique among Mammaliaformes but reminiscent of the condition in some derived tritylodontids11.
The lateral and ventral walls of the braincase also reveal a plethora of unusual features (Extended Data Fig. 2c, d), including an alisphenoid that is much larger than the anterior lamina of the petrosal, a robust and compact orbitosphenoid, a large bulbous ectopterygoid process (for possible attachment of the lateral pterygoid muscle) anterior to foramen ovale, the lack of fusion between presphenoid and basisphenoid (despite the advanced age of the individual), a cavum epiptericum that is almost completely floored ventrally, a cavum supracochleare (for the geniculate ganglion of cranial nerve VII) that is separate from the cavum epiptericum (for the semilunar ganglion of cranial nerve V), preservation of a possible process of the ossified pilae antotica and metoptica, and a single foramen (foramen ovale) for the mandibular division of the trigeminal nerve (cranial nerve V3) between the alisphenoid and the anterior lamina of the petrosal (as in non-mammaliaform cynodonts Megazostrodon and Haldanodon) but also possibly bordered by the pterygoid (unlike any known Mesozoic mammaliaform).
The basicranium of Vintana is also unique among Mesozoic mammaliamorphs in lacking a functional prootic canal and unusual in lacking a channel for the inferior petrosal sinus and any substantial branches of the stapedial system in the middle ear but in possessing an hiatus Fallopii that opens endocranially deep within the posterior wall of the cavum epiptericum (rather than in the lateral trough or at the anterior end of the petrosal), a tympanohyal that abuts the promontorium (as in monotremes), and a fossa incudis that is narrower than the epitympanic recess (as in therians) (Extended Data Figs 2c, 4). Vintana is also primitive in retaining a basioccipital lappet on the cochlear housing, as in the mammaliamorphs Adelobasileus and Sinoconodon. In the occipital region of the cranium, Vintana is the only Mesozoic mammaliaform for which both a postparietal and paired tabulars, together comprising the interparietal, have been identified as discrete elements (Fig. 1e; Extended Data Figs 2a, c, 5). However, the presence of these elements may be more commonplace among Mesozoic mammaliaforms, as recently revealed for extant mammals12. Finally, many of the cranial elements of Vintana contain a remarkable amount of cancellous bone (diploë) compared to those of other Mesozoic mammaliaforms (Extended Data Figs 2a, c and 3a), although a substantial amount is present in the basicranium of the much smaller Haldanodon13.
Endocranial and inner ear features
The mosaicism of derived and primitive features in Vintana is also exhibited in its endocranial morphology, a digital reconstruction of which (Extended Data Fig. 3a) reveals that the brain was small and similar in relative size to those of basal mammaliaforms (encephalization quotient = 0.28–0.56). The olfactory bulbs were very large, occupying over 14% of endocranial volume. Unlike the condition in other Mesozoic mammaliaforms, the endocast is strongly flexed (∼32° between the olfactory bulbs and the post-olfactory endocast). The cochlear canal, part of the osseous labyrinth of the inner ear, is only slightly curved and short (5.39 mm), only about half the length of the promontorium (Extended Data Fig. 3b). In these regards, Vintana resembles various non-mammaliaform cynodonts including tritylodontids, tritheledontids and Sinoconodon. By contrast, the presence of both primary and secondary osseous laminae, a tractus foraminosus, and Rosenthal’s canal represent much more derived characteristics of the mammaliaform inner ear.
Palaeobiology
Vintana sertichi is the largest known mammaliaform from the Mesozoic of Gondwana, superseded in Laurasia only by the eutriconodontan Repenomamus giganticus from the Early Cretaceous of China14. Estimated from the length of the cranium (124.1 mm long), the body mass of V. sertichi was 8.95 kg (95% confidence interval = 5.59–14.32 kg) (see Supplementary Information).
The direction of wear striations, orientation of enamel islets and synclines, and distribution of leading and trailing edges on the molariform teeth of Vintana indicate that the direction of the power stroke of the chewing cycle was primarily palinal (distal) (Extended Data Fig. 6), as in haramiyidans15, multituberculates16,17 and other gondwanatherians18,19. However, Vintana appears to be unique among these clades in possessing a significant buccal component to the power stroke. This distobuccal direction is corroborated by biomechanical analyses of the moments generated by the reconstructed primary jaw adductors around the dentary–squamosal joint axis (Extended Data Fig. 7; Supplementary Information). These analyses predict that Vintana had significantly higher bite forces than the similarly sized extant rodent Myocastor (Supplementary Information). Based on its large size, hypsodont molars and inferred relatively high bite forces, it is likely that Vintana had a mixed diet that included large, hard and/or abrasive food items such as roots, seeds or nut-like fruits, not unlike the abrasive, herbivorous diet inferred for other sudamericids4,5,6.
When compared with a range of extant mammals, Vintana had very large orbits (30–32 mm in diameter) relative to cranial size (Extended Data Fig. 8a). Although orbital size tends to progressively overestimate eye size as body mass and orbital diameter increase20,21,22, it is likely that Vintana also had relatively large eyes, as is the case for various extant felids, bovids and cervids23,24 (Extended Data Fig. 8a). Such large eyes could be consistent with either increased sensitivity under low light conditions or enhanced acuity across a range of ambient light levels, depending on eye structure. Furthermore, the radii of curvature of the semicircular canals in Vintana are very large (mean = 2.86 mm) for its estimated body mass (Extended Data Fig. 8b). Among living mammals, large semicircular canal radii of curvature are generally associated with large eye size25. The semicircular canals of Vintana are also almost mutually orthogonal, with angles between the canals ranging from 91° to 94°. This configuration represents only minor deviation from 90° compared to that exhibited by most other Mesozoic synapsids (for example, angles between the anterior and posterior semicircular canals range from 102° to 157° in non-mammaliaform synapsids26 and 65° to 80° in multituberculates27, but between 80° and 105° in Cretaceous eutherians28). The large size and orthogonality of the semicircular canals suggest that Vintana had high vestibular sensitivity to angular head accelerations29,30. These vestibular features may have evolved in order to stabilize large eyes during rapid and/or agile locomotion.
The inner ear of Vintana also exhibits cochlear primary and secondary osseous laminae and a cochlear canal that is relatively longer than in non-mammaliaform cynodonts but shorter than in extant therians (Extended Data Figs 3b, 8c). These cochlear features are probably associated with the presence of a stiff and short basilar membrane31, suggesting that Vintana had some capacity for high frequency hearing (that is, higher than 20 kilohertz) but that its cochlea may have encoded a more limited range of frequencies than the cochleae of most extant therians.
Finally, Vintana had very large olfactory bulbs relative to both its estimated body mass (Extended Data Fig. 8d) and endocranial volume (>14%). In this regard, it resembles some of the most basal mammaliaforms (for example, Morganucodon, Hadrocodium, Triconodon). Because the size of olfactory bulbs likely varies as a function of the number and size of the constituent glomeruli, which receive input from olfactory receptor neurons, it is reasonable to conclude that Vintana resembled many extant therian mammals in having an expanded olfactory receptor gene complement and in being able to detect and discriminate among a large number of odorant types32,33. These comparative data on the sensory anatomy of Vintana indicate that it possessed a distinctive suite of sensory adaptations compared to most other Mesozoic mammaliaforms, including large eyes, some capacity for high frequency hearing, and a keen sense of smell.
Phylogenetic relationships
Gondwanatherians have been variously regarded as Paratheria34, Xenarthra35, Multituberculata6,19, the sister-group to Multituberculata6,36,37, and Mammalia incertae sedis2,38. To assess the relationships of Gondwanatheria to other mammaliaforms, and generic interrelationships within Gondwanatheria, we undertook both parsimony and Bayesian phylogenetic analyses of 87 cynodont taxa (mostly Mesozoic mammaliaforms). This work builds upon previous data sets (see Supplementary Information) by modifying previously used characters, adding several new characters, and scoring several non-therian taxa not incorporated previously (see Supplementary Information for more detailed explanation of data, analysis and results).
Our results indicate that Gondwanatheria are monophyletic, composed primarily of the Sudamericidae, of which Vintana is a member, as is the previously unassigned Greniodon3 (Fig. 3). In all analyses Gondwanatheria are placed within the monophyletic Allotheria, including Haramiyavia, Thomasia, Arboroharamiya and Multituberculata (Figs 3, Supplementary Figs 1–4). Relationships among these clades, however, differ between different analytical approaches (see Supplementary Information). The clade containing Gondwanatheria is sister to Multituberculata in the parsimony analysis (Fig. 3, Supplementary Fig. 1), whereas they are nested within the latter in the Bayesian analysis (Supplementary Fig. 2). If this is indeed reflective of the true history of these clades, then several features generally accepted as plesiomorphic within Mammaliamorpha must have re-evolved in the lineage represented by Vintana (for example, basioccipital wing overlapping the cochlear housing, large septomaxilla with intranarial process, single trigeminal foramen between anterior lamina and alisphenoid).

Simplified strict consensus tree of 40 equally parsimonious trees (tree length 2,025; consistency index (CI) = 0.305; retention index (RI) = 0.701) derived from analysis of 453 characters and 87 cynodont taxa, with multistate characters unordered and unweighted. Thin black lines represent phylogenetic relationships and thick coloured lines indicate temporal ranges of taxa, with colours indicating supercontinent provenance. Vintana is highlighted in red font. Australosphenida includes Teinolophos, Steropodon, Bishops, Ausktribosphenos and Monotremata. Allotheria are highlighted in blue. Palaeogeographic maps of Gondwanan supercontinent modified from refs 40 (John Wiley and Sons), 42 and 50 (Elsevier). Madagascar highlighted in red to indicate its progressive geographic isolation through time. See Supplementary Information for taxon and character lists, data matrix, limitations and assumptions, phylogenetic methods, and more detailed explanation of results.
Mosaicism and evolution in isolation
Gondwanatherians are a strictly Gondwanan radiation, whereas multituberculates, their closest relatives, are overwhelmingly Laurasian in distribution2,10,39 (Fig. 3). The early evolution of gondwanatherians remains a mystery that can only be resolved with the discovery of more specimens from the tectonically most active interval of Gondwanan breakup (Middle Jurassic–Early Cretaceous). However, knowledge of the palaeogeographic history of Gondwana provides some insights concerning the lineage to which Vintana belongs. Madagascar, together with the Indian subcontinent, separated from Africa approximately 165 million years ago (Myr ago) and became fully isolated from Antarctica and Australia approximately 115–112 Myr ago, with Madagascar and the Indian subcontinent separating from each other about 88 Myr ago40,41,42 (Fig. 3, bottom). The basal stock that ultimately led to Vintana, at 72–66 Myr ago, was therefore likely isolated on Indo-Madagascar for about 24–27 million years and on Madagascar alone for another approximately 16–22 million years, for a total duration of 40 to almost 50 million years. Interestingly, support for the isolation of the Indo-Malagasy gondwanatherians can be derived from the Bayesian phylogenetic analyses, which identify a node (Vintana+Lavanify+Bharattherium) exclusive of the South American and African forms (see Supplementary Information).
The long period of geographic isolation of Indo-Madagascar and then Madagascar resulted in a latest Cretaceous Malagasy fauna that included a range of other unusual taxa (for example, massive predatory frogs43, herbivorous crocodyliforms44, and variously specialized theropod dinosaurs45,46,47). The ghost lineages of these taxa are long and indicate minimum divergence times near the Early–Late Cretaceous boundary (100 Myr ago) or even earlier40. Similarly, molecular divergence dates indicate early origins on Madagascar for xenotyphloid blind snakes, oplurid lizards and podocnemidid turtles41,48,49. We hypothesize that these and other lineages were more broadly distributed across Gondwana before fragmentation of the supercontinent, but became isolated on Indo-Madagascar when it separated from first Africa and then Antarctica and Australia. In this spatiotemporal context, Vintana appears to have retained several features reflective of its ancestry before early isolation from other Gondwanan landmasses on Indo-Madagascar and then Madagascar alone, but acquired its highly derived, unique morphology during 40–50 million years of evolution in isolation.
References
Rogers, R. R. et al. A new, richly fossiliferous member comprised of tidal deposits in the Upper Cretaceous Maevarano Formation, northwestern Madagascar. Cretac. Res. 44, 12–29 (2013)
Kielan-Jaworowska, Z., Cifelli, R. L. & Luo, Z.-X. Mammals From the Age of Dinosaurs: Origins, Evolution, and Structure (Columbia Univ. Press, 2004)
Goin, F. J. et al. Persistence of a Mesozoic, non-therian mammalian lineage (Gondwanatheria) in the mid-Paleogene of Patagonia. Naturwissenschaften 99, 449–463 (2012)
v. Koenigswald, W., Goin, F. & Pascual, R. Hypsodonty and enamel microstructure in the Paleocene gondwanatherian mammal Sudamerica ameghinoi. Acta Palaeontol. Pol. 44, 263–300 (1999)
Gurovich, Y. Additional specimens of sudamericid (Gondwanatheria) mammals from the early Paleocene of Argentina. Palaeontology 51, 1069–1089 (2008)
Gurovich, Y. & Beck, R. The phylogenetic affinities of the enigmatic mammalian clade Gondwanatheria. J. Mamm. Evol. 16, 25–49 (2009)
Rougier, G. W. Vincelestes neuquenianus Bonaparte (Mammalia, Theria) un Primitivo Mamífero del Cretácico Inferior de la Cuenca Neuquina. PhD thesis, Univ. Buenos Aires. (1993)
Rougier, G. W., Apestiguía, S. & Gaetano, L. C. Highly specialized mammalian skulls from the Late Cretaceous of South America. Nature 479, 98–102 (2011)
Krause, D. W., Prasad, G. V. R., von Koenigswald, W., Sahni, A. & Grine, F. E. Cosmopolitanism among Late Cretaceous Gondwanan mammals. Nature 390, 504–507 (1997)
Krause, D. W. Gondwanatheria and ?Multituberculata (Mammalia) from the Late Cretaceous of Madagascar. Can. J. Earth Sci. 50, 324–340 (2013)
Clark, J. M. & Hopson, J. A. Distinctive mammal-like reptile from Mexico and its bearing on the phylogeny of the Tritylodontidae. Nature 315, 398–400 (1985)
Koyabu, D., Maier, W. & Sánchez-Villagra, M. R. Paleontological and developmental evidence resolve the homology and dual embryonic origin of a mammalian skull bone, the interparietal. Proc. Natl Acad. Sci. USA 109, 14075–14080 (2012)
Ruf, I., Luo, Z.-X. & Martin, T. Re-investigation of the basicranium of Haldanodon exspectatus (Docodonta, Mammaliaformes). J. Vertebr. Paleontol. 33, 382–400 (2013)
Hu, Y., Meng, J., Wang, Y. & Li, C. Large Mesozoic mammals fed on young dinosaurs. Nature 433, 149–152 (2005)
Butler, P. M. Review of the early allotherian mammals. Acta Palaeontol. Pol. 45, 317–342 (2000)
Krause, D. W. Jaw movement, dental function, and diet in the Paleocene multituberculate Ptilodus. Paleobiology 8, 265–281 (1982)
Gambaryan, P. P. & Kielan-Jaworowska, Z. Masticatory musculature of Asian taeniolabidoid multituberculate mammals. Acta Palaeontol. Pol. 40, 45–108 (1995)
Krause, D. W., Kielan-Jaworowska, Z. & Bonaparte, J. F. Ferugliotherium Bonaparte, the first known multituberculate from South America. J. Vertebr. Paleontol. 12, 351–376 (1992)
Krause, D. W. & Bonaparte, J. F. Superfamily Gondwanatherioidea: a previously unrecognized radiation of multituberculate mammals in South America. Proc. Natl Acad. Sci. USA 90, 9379–9383 (1993)
Schultz, A. H. The size of the orbit and of the eye in primates. Am. J. Phys. Anthropol. 26, 389–408 (1940)
Kay, R. F. & Kirk, E. C. Osteological evidence for the evolution of activity pattern and visual acuity in primates. Am. J. Phys. Anthropol. 113, 235–262 (2000)
Kirk, E. C. Effects of activity pattern on eye size and orbital aperture size in primates. J. Hum. Evol. 51, 159–170 (2006)
Ritland, S. M. The Allometry of the Vertebrate Eye PhD thesis, Univ. Chicago (1982)
Ross, C. F. & Kirk, E. C. Evolution of eye size and shape in primates. J. Hum. Evol. 52, 294–313 (2007)
Kemp, A. D. & Kirk, E. C. Eye size and visual acuity influence vestibular anatomy in mammals. Anat. Rec. 297, 781–790 (2014)
Olson, E. C. Origin of mammals based upon the cranial morphology of therapsid suborders. Spec. Pap. Geol. Soc. Am. 55, 1–130 (1944)
Hurum, J. H. The inner ear of two Late Cretaceous multituberculate mammals, and its implications for multituberculate hearing. J. Mamm. Evol. 5, 65–93 (1998)
Ekdale, E. G. Comparative anatomy of the bony labyrinth (inner ear) of placental mammals. PLoS ONE 8, e66624 (2013)
Yang, A. & Hullar, T. E. Relationship of semicircular canal size to vestibular-nerve afferent sensitivity in mammals. J. Neurophysiol. 98, 3197–3205 (2007)
Berlin, J. C., Kirk, E. C. & Rowe, T. B. Functional implications of ubiquitous semicircular canal non-orthogonality in mammals. PLoS ONE 8, e79585 (2013)
Luo, Z. X., Ruf, I. & Martin, T. The petrosal and inner ear of the Late Jurassic cladotherian mammal Dryolestes leiriensis and implications for ear evolution in therian mammals. Zool. J. Linn. Soc. 166, 433–463 (2012)
Mori, K., Nagao, H. & Yoshihara, Y. The olfactory bulb: coding and processing of odor molecule information. Science 286, 711–715 (1999)
Niimura, Y. Olfactory receptor multigene family in vertebrates: from the viewpoint of evolutionary genomics. Curr. Genomics 13, 103–114 (2012)
Scillato-Yané, G. J. & Pascual, R. Un peculiar Paratheria, Edentata (Mammalia) del Paleoceno de Patagonia. Primeras J. Argent. Paleontol. Vertebr., abstr. 15. (1984)
Scillato-Yané, G. J. & Pascual, R. Un peculiar Xenarthra del Paleoceno medio de Patagonia (Argentina). Su importancia en la sistemática de los Paratheria. Ameghiniana 21, 173–176 (1985)
Gurovich, Y. Bio-evolutionary Aspects of Mesozoic Mammals: Description, Phylogenetic Relationships and Evolution of the Gondwanatheria (Late Cretaceous and Paleocene of Gondwana) PhD thesis, Univ. Buenos Aires (2006)
Pascual, R. & Ortiz-Jaureguizar, E. The Gondwanan and South American episodes: two major and unrelated moments in the history of the South American mammals. J. Mamm. Evol. 14, 75–137 (2007)
Pascual, R., Goin, F. J., Krause, D. W., Ortiz-Jaureguizar, E. & Carlini, A. A. The first gnathic remains of Sudamerica: implications for gondwanathere relationships. J. Vertebr. Paleontol. 19, 373–382 (1999)
Rich, T. H. et al. An Australian multituberculate and its palaeobiogeographic implications. Acta Palaeontol. Pol. 54, 1–6 (2009)
Ali, J. R. & Krause, D. W. Late Cretaceous bioconnections between Indo-Madagascar and Antarctica: refutation of the Gunnerus Ridge causeway hypothesis. J. Biogeogr. 38, 1855–1872 (2011)
Samonds, K. E. et al. Spatial and temporal arrival patterns of Madagascar’s vertebrate fauna explained by distance, ocean currents, and ancestor type. Proc. Natl Acad. Sci. USA 109, 5352–5357 (2012)
Samonds, K. E. et al. Imperfect isolation: factors and filters shaping Madagascar’s extant vertebrate fauna. PLoS ONE 8, e62086 (2013)
Evans, S. E., Jones, M. E. H. & Krause, D. W. A giant frog with South American affinities from the Late Cretaceous of Madagascar. Proc. Natl Acad. Sci. USA 105, 2951–2956 (2008)
Buckley, G. A., Brochu, C., Krause, D. W. & Pol, D. A pug-nosed crocodyliform from the Late Cretaceous of Madagascar. Nature 405, 941–944 (2000)
Forster, C. A., Sampson, S. D., Chiappe, L. M. & Krause, D. W. The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar. Science 279, 1915–1919 (1998)
Sampson, S. D. et al. Predatory dinosaur remains from Madagascar: implications for the Cretaceous biogeography of Gondwana. Science 280, 1048–1051 (1998)
Sampson, S. D., Carrano, M. T. & Forster, C. A. A bizarre predatory dinosaur from the Late Cretaceous of Madagascar. Nature 409, 504–506 (2001)
Crottini, A. et al. Vertebrate time-tree elucidates the biogeographic pattern of major biotic change around the K-T boundary in Madagascar. Proc. Natl Acad. Sci. USA 109, 5358–5363 (2012)
Vidal, N. et al. Blindsnake evolutionary tree reveals long history on Gondwana. Biol. Lett. 6, 558–561 (2010)
Ali, J. R. & Aitchison, J. C. Gondwana to Asia: plate tectonics, paleogeography and the biological connectivity of the Indian sub-continent from the Middle Jurassic through latest Eocene (166–35 Ma). Earth Sci. Rev. 88, 145–166 (2008)
Acknowledgements
We thank the Université d’Antananarivo, the Madagascar Institut pour la Conservation des Ecosystèmes Tropicaux, and the villagers of the Lac Kinkony Study Area for logistical support of fieldwork; various ministries of the Republic of Madagascar for permission to conduct field research; members of the 2010 field research team for their efforts; J. Thostenson and M. Hill of the American Museum of Natural History Microscopy & Imaging Facility, New York, New York, and J. Diehm and B. Ruether of Avonix Imaging, Plymouth, Minnesota, and various members of the Department of Radiology at Stony Brook University for providing expert assistance in computed tomography scanning; L. Betti-Nash for artwork; J. Neville for photography; D. Pulaski for reconstructing the cranium and building the finite element models; Z.-X. Luo, G. Rougier and A. Weil for their reviews of the paper; and the National Geographic Society (8597-09) and the National Science Foundation (EAR-0446488, EAR-1123642) for funding.
Author information
Authors and Affiliations
Contributions
J.R.G. prepared the fossil; J.R.G., S.H., W.L.H. and P.M.O. conducted most of the micro-computed tomography digital preparation; L.J.R. and R.R.R. provided geological data; H.A. provided logistical support; J.R.G., S.H., D.W.K., W.v.K., P.M.O., J.B.R. and J.R.W. provided most of the descriptions and comparisons; E.R.D., A.D.K., D.W.K., E.C.K., W.v.K., P.M.O., J.A.S. and J.R.W. conducted various functional and comparative analyses; S.H., D.W.K., E.R.S. and J.R.W. contributed to the phylogenetic analysis; D.W.K. developed the manuscript, with contributions from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Vintana sertichi has been assigned the Life Science Identifier (LSID) http://zoobank.org/urn:lsid:zoobank.org:act:B21CC5B2-D550-4D78-BA1F-8319EA663785.
Extended data figures and tables
Extended Data Figure 1 Enamel microstructure of Vintana sertichi.
Scanning electron micrograph of transverse section of enamel (sampled from distolingual corner of left MF2) revealing the radial arrangement of prisms (p) separated by well-developed interrow sheets (irs) of interprismatic matrix.
Extended Data Figure 2 Craniofacial and endocranial morphology of Vintana sertichi.
a, Reconstruction of right half of cranium (UA 9972) in medial view showing bony composition; sagittally sectioned elements shaded in grey, endocranial cavity shaded in blue. b, Reconstruction of right half of anterior portion of cranium in medial view showing details of nasal cavity, anterior part of endocranium, and nasopharyngeal canal. Dorsoventral extent of cribriform plate indicated in red. Paranasal lamella of nasoturbinal ridge indicated by six short red arrows pointing to a ridge ventral to tectal lamella. c, Digital reconstruction of mid-sagittal cut-away medial view of right posterior half of cranium based on micro-computed tomography scans. d, Digital reconstruction of lateral view of right posterior half of cranium, based on micro-computed tomography scans (squamosal cut away ventrally, indicated by hatching; basisphenoid and other more medial elements removed to show openings). AS, alisphenoid; BS, basisphenoid; ce, cavum epiptericum; cpl, cribriform plate ridge; cs, crista semicircularis; ep, ectopterygoid process; ethr, ethmoturbinal ridges; fo, foramen ovale; FR, frontal; hf, hypoglossal foramina; iam, internal acoustic meatus; IP, interparietal; jf, jugular foramen; LA, lacrimal; lret 1, lateral root of ethmoturbinal 1; MX, maxilla; nlcg, nasolacrimal canal and groove; nvg, neurovascular grooves; OC, occipital; occ, occipital condyle; opf, optic foramen; opg, optic groove; OS, orbitosphenoid; PA, parietal; PAL, palatine; ?pan, possible pila antotica; PE, petrosal; pmet, pila metoptica; ppr, paroccipital process; pr, promontorium; prof, prootic foramen; PT, pterygoid; ptf, posttemporal foramen; ptl, posterior transverse lamina; RMF3, right third upper molariform tooth; rt, foramen for ramus temporalis; saf, subarcuate fossa; sof, sphenorbital fissure; SQ, squamosal; tlnt, tectal lamella of nasoturbinal ridge; vret 1, vertical root of ethmoturbinal 1.
Extended Data Figure 3 Endocranial and inner ear morphology of Vintana sertichi.
a, Digital reconstruction of mid-sagittal cut-away view of right half of cranium and integrated endocranial cast reconstruction, based on micro-computed tomography scans, to reveal position and orientation of endocast (blue; moderately transparent), right inner ear (magenta; petrosal is rendered as fully transparent), and positions of right cranial nerves (CNs, green). b, Digital reconstruction of right osseous labyrinth and cochlear nerve (cranial nerve VIII), based on micro-computed tomography scans, in medial view. Virtual endocast of osseous labyrinth (translucent), with canal system occupied by cochlear nerve (green) depicted both in situ (lower right) and in identical view but isolated and enlarged (upper left). asc, anterior semicircular canal; cc, crus commune; cn, cochlear nerve (part of cranial nerve VIII); “CN” II, optic nerve; CN V3, mandibular division of trigeminal nerve; CN VII, facial nerve; CN VIII, vestibulocochlear nerve; CN IX, glossopharyngeal nerve; CN X, vagus nerve; CN XII, hypoglossal nerve bundles (XIIa and XIIb); hp, habenulae perforatae; lsc, lateral semicircular canal; pf, perilymphatic foramen; psc, posterior semicircular canal; Rc, Rosenthal’s canal (cochlear ganglion canal); scc, secondary crus commune; sl, secondary osseous lamina of cochlea; tf, tractus foraminosus.
Extended Data Figure 4 Composition and features of basicranium of Vintana sertichi.
a, Ventral view of UA 9972. b, unlabelled, and b′, labelled views of basicranial region indicated by red rectangle in a. Occipital shaded in transparent blue, petrosal in yellow, and squamosal in orange. AS, alisphenoid; BO, basioccipital; BS, basisphenoid; cp, crista parotica; eam, external acoustic meatus; entpt, entopterygoid process; EO, exoccipital; er, epitympanic recess; fi, fossa incudis; fm, foramen magnum; fsh, facet for stylohyal; gf, glenoid fossa; hf, hypoglossal foramina; jf, jugular foramen; lc, fossa for longus capitis; lfl, lateral flange; lt, lateral trough; mrb, median ridge of basioccipital; mt, muscular tubercle; occ, occipital condyle; PE, petrosal; ppr, paroccipital process; ppts, post-promontorial tympanic sinus; pr, promontorium; PT, pterygoid; ptr, pterygopalatine ridge; rcv, fossa for rectus capitis ventralis; sf, stapedius fossa; smn, stylomastoid notch; SQ, squamosal; th, tympanohyal.
Extended Data Figure 5 Composition and features of occipital region of Vintana sertichi.
Posterior view of UA 9972. fm, foramen magnum; FR, frontal; jf, jugal flange; JU, jugal; nc, nuchal crest; OC, occipital; occ, occipital condyle; PA, parietal; PE, petrosal; pgs, postglenoid shelf; PP, postparietal; ppr, paroccipital process; PT, pterygoid; ptf, posttemporal foramen; SQ, squamosal; TA, tabular; zpj, zygomatic process of jugal; zpsq, zygomatic process of squamosal.
Extended Data Figure 6 Dental wear features of Vintana sertichi.
a, Scanning electron micrograph of oblique view of buccal side of occlusal surface of left MF4 cast (see Fig. 2c, d) showing leading (le) and trailing (tr) edges of enamel ridges; white arrow is parallel to wear striae and indicates distobuccal direction of movement of antagonistic lower molariforms. b, Rose diagram of measured wear striation directions on left MF4; black arrow represents mean vector angle of all striations.
Extended Data Figure 7 Relative contributions of the primary muscles of mastication to the total moments about the dentary-squamosal joint axis (DSJ).
Contributions illustrated by vector length during molariform/molar biting at a, b, narrow; and c, d, wide gapes in Vintana sertichi (left) and Myocastor coypus (right). For comparative purposes, the crania are shown at the same length. Only muscles that contributed more than 2% to the total moment about the DSJ axis are illustrated here. This represents 96–98% of the total moment about the axis. adm, anterior deep masseter; iozm, infraorbital portion of zygomaticomandibularis; sm, superficial masseter; t, temporalis; zm, zygomaticomandibularis. All muscles except zm (shown in red) attach to the lateral surface of the cranium and dentary.
Extended Data Figure 8 Sensory anatomy and ecology of Vintana sertichi.
a, Relationship between cranial size and orbital diameter in 352 extant mammalian species (identified according to order; see inset) and V. sertichi (both upper (32 mm) and lower (30 mm) estimates of orbital diameter are depicted). 1, 2, 3, bovids Oreotragus oreotragus, Ourebia ourebi and Raphicerus campestris; 4, 5, 6, felids Lynx rufus, Leptailurus serval and Leopardus pardalis; 7, cervid Mazama gouazoubira. b, Relationship between mean semicircular canal radius of curvature and body mass in 205 extant mammalian species and V. sertichi. The three symbols for Vintana represent its lower, middle and upper body mass estimates. c, Relationship between cochlear canal length and body mass in extinct and extant synapsids, including V. sertichi. The three symbols for Vintana represent its lower, middle and upper body mass estimates. The grey polygon encompasses extant therian mammals. The values for living monotremes do not include the lagenar portion (1, Tachyglossus; 2, Ornithorhynchus). d, Relationship between olfactory bulb volume and body mass in 163 extant and extinct Mammaliaformes, including V. sertichi depicted at lower, average and upper body mass estimates for the reconstructed olfactory bulb volume. See Supplementary Information for more detailed explanations and sources of data.
Supplementary information
Supplementary Information
This file contains Supplementary Text and Data – see contents page for details. (PDF 5365 kb)
Virtual reconstruction of cranium of Vintana sertichi (UA 9972) from μCT dataset, with full rotation (360°) about a dorsoventral axis.
Visualization maps dataset density contrast with false colors resembling those on actual specimen, with the exception of tooth enamel. (MOV 20234 kb)
Virtual reconstruction of cranium of Vintana sertichi (UA 9972) from µCT dataset, with full rotation (360°) about an anteroposterior axis.
Visualization maps dataset density contrast with false colors resembling those on actual specimen, with the exception of tooth enamel. (MOV 21651 kb)
Virtual reconstruction of elements of right side of cranium of Vintana sertichi (UA 9972).
Animation of translation of cranial elements from articulated position to Beauchene-style ("exploded") presentation. Cranial elements presented as polygon surfaces created from segmentation of μCT dataset. (MOV 21674 kb)
Virtual reconstruction of left molariform toothrow (MF2–MF4) of Vintana sertichi (UA 9972) from µCT dataset, with two full rotations (each 360°) about an anteroposterior axis.
First rotation highlights surface morphology. Second rotation highlights internal morphology, with pulp cavities in green and infundibula in blue. (MOV 20924 kb)
Virtual reconstruction of left molariform toothrow (MF2–MF4) of Vintana sertichi (UA 9972) from µCT dataset, with two full rotations (each 360°) about a dorsoventral axis.
First rotation highlights surface morphology. Second rotation highlights internal morphology, with pulp cavities in green and infundibula in blue. (MOV 16652 kb)
Rights and permissions
About this article

Cite this article
Krause, D., Hoffmann, S., Wible, J. et al. First cranial remains of a gondwanatherian mammal reveal remarkable mosaicism. Nature 515, 512–517 (2014). https://doi.org/10.1038/nature13922
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature13922
This article is cited by
-
Derived faunivores are the forerunners of major synapsid radiations
Nature Ecology & Evolution (2023)
-
New Skull Material of Taeniolabis taoensis (Multituberculata, Taeniolabididae) from the Early Paleocene (Danian) of the Denver Basin, Colorado
Journal of Mammalian Evolution (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
