
Abstract
Integrin-mediated adhesion regulates membrane binding sites for Rac1 within lipid rafts. Detachment of cells from the substratum triggers the clearance of rafts from the plasma membrane through caveolin-dependent internalization. The small GTPase Arf6 and microtubules also regulate Rac-dependent cell spreading and migration, but the mechanisms are poorly understood. Here we show that endocytosis of rafts after detachment requires F-actin, followed by microtubule-dependent trafficking to recycling endosomes. When cells are replated on fibronectin, rafts exit from recycling endosomes in an Arf6-dependent manner and return to the plasma membrane along microtubules. Both of these steps are required for the plasma membrane targeting of Rac1 and for its activation. These data therefore define a new membrane raft trafficking pathway that is crucial for anchorage-dependent signalling.
Similar content being viewed by others
Main
The small GTPases Rac, Rho and Cdc42 localize to ordered plasma membrane microdomains called lipid rafts1,2,3,4,5. These regions are enriched in cholesterol, sphingolipids, glycosylphosphatidylinositol (GPI)-linked proteins and other proteins6. Although some aspects of raft biology have been controversial, a consensus is emerging that they are small, dynamic structures whose properties are strongly influenced by their protein content7. They can modulate many signalling pathways in diverse biological processes such as cell division, apoptosis, adhesion and chemotaxis6. Lipid rafts modulate the spatial targeting of GTPases that is required for cell spreading and migration1,8. Interestingly, adhesion sites have very high membrane order, suggesting a raft-like composition9.
In anchorage-dependent cells, loss of integrin-mediated adhesion triggers the rapid and efficient endocytosis of multiple raft components including ganglioside GM1, GPI-linked proteins and cholesterol, leading to markedly decreased plasma membrane order1,9. Rafts are endocytosed through caveolae in a process that requires dynamin-2 and the phosphorylation of a small fraction of caveolin-1 on tyrosine 14 (pY14Cav1)2. The resultant loss of anchoring sites in the plasma membrane downregulates Rac1, ERK (extracellular signal-regulated kinase) and phosphatidylinositol-3-OH kinase-dependent pathways. Integrin-specific readhesion triggers the trafficking of raft components back to the plasma membrane and restores anchorage-dependent signalling.
The small GTPase Arf6 regulates endocytosis, post-endocytic recycling, exocytosis and cytoskeletal organization10,11. Importantly, Arf6 and several of its regulators are commonly overexpressed in metastatic cancers and are strongly implicated in the control of cell migration and spreading12. Consistent with this role is the observation that activation of Arf6 leads to increased GTP loading of Rac1 and movement of Rac1 to the plasma membrane13.
Microtubules also modulate the activity and function of Rac, as well as cell spreading and migration14,15. They do so in part by serving as tracks for intracellular vesicular trafficking. In addition, MTs directly bind regulators of Rho-family GTPases such as GEFH1 (Rho/Rac guanine nucleotide exchange factor 1)16.
In this study, we further examined the adhesion-dependent endocytosis and exocytosis of raft components in fibroblasts using a well-characterized lipid raft marker, cholera toxin subunit B (CTxB), which binds ganglioside mannoside 1 (GM1). Our results identify key components of this trafficking pathway and unexpectedly reveal the mechanisms by which microtubules and Arf6 regulate cell spreading.
Results
Cytoskeletal regulation of raft endocytosis
We first tested the role of F-actin and microtubules in the caveolin-dependent endocytosis of rafts in mouse embryo fibroblasts (MEFs) after their detachment from the tissue-culture plastic substratum on which they adhere to a matrix composed mainly of fibronectin. Cells were prelabelled with CTxB while still adherent. Some cells were first pretreated with the actin-depolymerizing drug latrunculin A (LatA; 1 μM) or the microtubule-depolymerizing agent nocodazole (10 μM), then detached and held in suspension for up to 90 min in the continued presence of drugs. In control cells, CTxB began to internalize immediately after detachment and moved to a distinct perinuclear location by 90 min (Fig. 1a). When CTxB was instead added 90 min after detachment, binding was minimal, indicating clearance of GM1 from the cell surface (Fig. 1b). Latrunculin A prevented the complete internalization of GM1 in suspension, indicated by the retention of CTxB surface labelling (Fig. 1b). F-actin is therefore required for the internalization of rafts through this pathway.

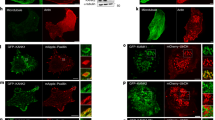
(a) Stably adherent MEFs pretreated with 1 μM latrunculin A (LatA), 10 μM nocodazole (NOC) or dimethylsulphoxide (control) were surface-labelled with CTxB-Alexa 488, then detached and held in suspension in the continued presence of drugs for 90 min. (b) Unlabelled cells suspended for 90 min were surface-labelled with CTxB-Alexa 594. Cells with no visible surface GM1 are marked with an asterisk. (c) Cells prelabelled with CTxB-Alexa 488 (as in a) (GM1-CTxB) were suspended for 90 min, then fixed and stained for γ-tubulin.
Nocodazole had no effect on detachment-induced internalization itself, as indicated by the loss of surface labelling by CTxB in suspended cells (Fig. 1b). However, in prelabelled cells, internalized CTxB remained in the cell periphery, failing to move to the cell centre (Fig. 1a). When prelabelled control cells were fixed and stained for γ-tubulin, perinuclear CTxB was localized around the microtubule organizing centre (MTOC; Fig. 1c; Pearson coefficient 0.72 ± 0.042). Thus, internalized CTxB seems to be transported towards the minus end of the microtubules, to the MTOC.
Identification of the intracellular compartment
We next sought to identify the intracellular compartment to which CTxB localizes in suspended cells. The proximity of the Golgi to the MTOC, together with known trafficking of CTxB to this compartment in adherent cells17 made it an obvious candidate. In suspended cells, endocytosed CTxB and the cis-Golgi marker GM130 seemed to colocalize (Fig. 2a). However, careful examination at higher resolution showed CTxB to be present in a distinct structure that was complementary to the GM130 stain (Fig. 2a; Pearson coefficient 0.20 ± 0.05, significantly less (P < 0.001) than the overlap seen with MTOC). This pattern could be seen most clearly when the intensity of the GM130 signal was gradually increased in merged images (Fig. 2a, lower panel).

(a) Stably adherent MEFS surface-labelled with CTxB-Alexa 594 (GM1-CTxB) were suspended for 90 min, then fixed and stained for the Golgi marker GM130 (GM130-FITC). The lower panel shows an enlarged view of the cell centre with the intensity of GM130 (green) increased stepwise and that of CTxB-Alexa 594 (red) held constant. (b) Suspended (90 min) MEFs with endocytosed CTxB-Alexa 594 (GM1-CTxB) were untreated (control) or treated with 10 μg ml−1 brefeldin A (+BFA) for 30 min. Cells were fixed and stained for GM130. (c) MEFs expressing GFP-tagged PKD-KD mutant (K618N) were surface labelled with CTxB-Alexa 594, detached for 90 min to allow endocytosis, then replated on fibronectin (10 μg ml−1) for 30 min. (d) MEFs surface labelled with CTxB-Alexa 594 (CTxB) and incubated for 90 min in suspension were stained for AMF-R.
To confirm this result, cells were treated with brefeldin A (BFA), which induces vesiculation and dispersion of the Golgi. Although BFA clearly dispersed the cis-Golgi marker GM130 (Fig. 2b) and the trans-Golgi marker TGN38 (data not shown), endocytosed CTxB was unaffected (Fig. 2b). Furthermore, BFA did not noticeably inhibit cell spreading or the return of CTxB to the cell surface in cells replated on fibronectin (Supplementary Information, Fig. 1a). Finally, a kinase-dead mutant of protein kinase D (PKD-KD; K618N) that blocks the exit of newly synthesized proteins from the Golgi18 also had no effect on the movement of CTxB from the perinuclear pool (Fig. 2c) to the cell surface in replated cells (Supplementary Information, Fig. 1c). It did, however, block transport of the temperature-sensitive vesicular stomatis viral G protein fused to green fluorescent protein (VSVG–GFP) protein19 from the Golgi (Supplementary Information, Fig. 1d), confirming its efficacy. A PKD-KD (P155/287G) mutant not localized to the Golgi was used as a control (Supplementary Information, Fig. 1b, d). VSVG–GFP localized in the Golgi also showed minimal overlap with endocytosed CTxB (Supplementary Information, Fig. 1e). Taken together, these data exclude the Golgi as the location of endocytosed GM1 in suspended cells.
SV40 internalizes through caveolae to the smooth endoplasmic reticulum (SER)20; we therefore examined this organelle. The SER marker autocrine motility factor receptor (AMF-R) showed no colocalization with endocytosed CTxB (Fig. 2d), suggesting that it is not trafficked to the SER.
Recycling endosomes localize near the MTOC and, in adherent cells, contain endocytosed rafts17,21. To test this location, we used the well-characterized recycling endosome marker Rab11 (ref. 22). Endocytosed CTxB in suspended cells showed a strong overlap with GFP–Rab11 (Fig. 3a; Pearson coefficient 0.72 ± 0.06, significantly higher (P < 0.001) than the overlap seen with GM130). Moreover, Rab11 localization and its overlap with CTxB were resistant to BFA (data not shown). We also examined internalized transferrin (Tf), which is efficiently trafficked to the recycling endosomes after clathrin-dependent uptake23. Adherent cells that had been surface-labelled with Tf-Alexa 594 were subsequently labelled with CTxB-Alexa 488 and detached. Endocytosed CTxB strongly colocalized with Tf-Alexa 594 at 30 min (Fig. 3b, lower panel; Pearson coefficient 0.75 ± 0.045, significantly higher (P < 0.001) than the overlap seen with GM130). The recycling endosomes are therefore the site of internalized GM1 in suspended cells.

(a) MEFs expressing Rab11–GFP were surface-labelled with CTxB-Alexa 594 (CTxB) and incubated in suspension for 90 min. The central region is enlarged in the rightmost panel. (b) Adherent MEFs labelled with transferrin-Alexa 594 (transferrin) for 30 min were detached and surface-labelled with CTxB-Alexa 488 on ice. Cells were fixed immediately (susp. 0 min) or warmed to 37 °C for 30 min (susp. 30 min). The central region is enlarged in the rightmost panel. (c) MEFs expressing Rab11(S25N)–GFP were surface labelled with CTxB-Alexa 594 (GM1-CTxB), held in suspension for 90 min and replated on 10 μg ml−1 fibronectin for 15 min. Readherent cells were labelled with transferrin-Alexa 594 (transferrin) for 60 min at 37 °C (lower panels). Non-transfected control cells are marked with asterisks. (d) MEFs expressing GFP-tagged WT Arf6 were surface-labelled with CTxB-Alexa 594 (GM1-CTxB) and incubated in suspension for 90 min. The central region is enlarged in the rightmost panel. (e) Cells expressing unlabelled GFP-tagged WT Arf6 suspended for 90 min were replated on fibronectin (10 μg ml−1) for 15 min and surface GM1 was labelled with CTxB-Alexa 594. Membrane ruffles are enlarged in the rightmost panel.
Rab11 regulates the trafficking of TfR from recycling endosomes to the plasma membrane22. To test whether it similarly regulates GM1, we examined the effects of dominant-negative S25N Rab11. Although S25N Rab11 blocked the exit of Tf from recycling endosomes (Fig. 3c, and Supplementary Information, Fig. 2b), it affected neither the return of CTxB to the cell surface after replating (Supplementary Information, Fig. 2a) nor cell spreading (Fig. 3c). In cells prelabelled with both CTxB-Alexa 488 and Tf-Alexa 594, these fluorophores showed little colocalization during cell spreading (Supplementary Information, Fig. 1f). Therefore, although both GM1 and Tf traffic from the recycling endosomes to the plasma membrane during readhesion, they do so by means of distinct pathways.
Rab22a also localizes to recycling endosomes and regulates the trafficking of membranes endocytosed by clathrin-independent pathways24. However, dominant-negative S19N Rab22 did not affect the endocytosis of GM1 after detachment, nor did it affect its exocytosis or the spreading of replated cells (Supplementary Information, Fig. 2c, d). Rab22a therefore does not seem to regulate this pathway.
Role of Arf6 in raft trafficking
Arf6 can regulate vesicular trafficking from recycling endosomes25. Intriguingly, Arf6 was also reported to control the movement of Rac1 from endosomes to the plasma membrane and Rac activation13,26. This similarity to integrin-mediated adhesion prompted us to test the role of Arf6 in raft trafficking. In suspended MEFs, GFP-tagged wild-type (WT) Arf6 strongly overlapped with endocytosed CTxB-Alexa 594 in recycling endosomes (Fig. 3d; Pearson coefficient 0.71 ± 0.004, significantly higher (P < 0.001) than the overlap seen with GM130). Surface CTxB in spreading cells also colocalized extensively with Arf6 at lamellipodia and membrane ruffles (Fig. 3e).
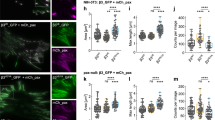
We next tested the effects of dominant-negative (T27N) Arf6, which blocks both cell spreading13,27 and the trafficking of some components out of recycling endosomes25. To determine whether effects on spreading were mediated by the trafficking of the components of lipid rafts, we examined MEFs from both caveolin-1−/− (Cav1−/−) mice and matched WT littermates. Unlike the WT, Cav1−/− MEFs do not internalize raft markers after detachment and retain functional, membrane-localized active Rac1 even in suspension1. Thus, spreading of Cav1−/− cells should not require the return of raft components to the plasma membrane. Cells transiently transfected with WT or T27N Arf6 were detached, held in suspension for 90 min and replated on fibronectin. T27N Arf6 inhibited the spreading of WT cells as expected but had absolutely no effect on Cav1−/− cells (Fig. 4a, b). T27N Arf6 also inhibited the appearance of GM1 on the cell surface in readherent WT cells but not Cav1−/− cells (Fig. 4b). However, at later times after replating (45 min or more), spreading and surface GM1 in T27N Arf6-expressing cells recovered (Supplementary Information, Fig. 2e, f). Thus, inhibiting Arf6 slows rather than completely blocks GM1 exocytosis and cell spreading. Inhibition by T27N Arf6 was not the result of changes in integrin levels or function, because the binding of cells to fibronectin was unaffected (Supplementary Information, Fig. 2g). WT Arf6 had no effect on spreading, surface GM1 levels or raft endocytosis compared with non-transfected cells (data not shown).

(a) Stably adherent WT and Cav1−/−; MEFs expressing HA-tagged WT Arf6 (WT) or T27N Arf6 were suspended for 90 min and replated on fibronectin (2 μg ml−1) for 15 min. Cells were then chilled and surface-labelled with CTxB-Alexa 594, and images were taken (top). Surface areas were measured (bottom); values are means and s.e.m. for n = 25 cells, in arbitrary units. The figure is representative of three independent experiments. (b) Cells suspended for 90 min were replated on fibronectin (2 μg ml−1) for 15 min. Suspended and replated cells were surface-labelled with CTxB and bound CTxB was detected by western blotting (CTxB), using tubulin as a loading control (tubulin). Numbers alongside the blots represent molecular weight markers. Bands were quantified (bottom); values (means and s.e.m., n = 3, normalized for tubulin) were calculated relative to WT MEFs expressing WT Arf6. (c) Cells transfected with a vector coding for both GFP and Arf6 shRNA (KD) or scrambled control RNA (con.) were suspended, replated on fibronectin (2 μg ml−1) for 15 min, and labelled with CTxB-Alexa 594 (left; as in a). Surface areas were quantified (top); values are means and s.e.m. for n = 55 cells, in arbitrary units. The figure is representative of two independent experiments. (d) Quantification of bound CTxB detected by western blotting. Values (means and s.e.m., n = 3; normalized for tubulin) were calculated relative to control WT MEFs. (e) WT and Cav1−/−; MEFs expressing HA-tagged WT Arf6 (WT) or T27N Arf6 were suspended for 90 min (susp. 90 min) and replated on fibronectin (2 μg ml−1) for 15 min (FN 15 min) or 45 min (FN 45 min). Cells were lysed and fractionated as described in Methods. Rac1 in the membrane fraction was determined as described in Methods. Values are means and s.e.m. (n = 3), in arbitrary units. (f) Active Rac1 was measured with the GST–PBD pulldown assay, and its percentage relative to total Rac1 was calculated. Values are means and s.e.m. (n = 3). Uncropped images of the scans in (b) are shown in Supplementary Information, Fig. 7.
Specific knockdown of Arf6 (Supplementary Information, Fig. 3d, e) also slowed cell spreading and the appearance of GM1 on the surface of WT but not Cav1−/− MEFs (Fig. 4c, d). Surface levels of GPI-linked proteins, which also localize to raft domains, were similarly regulated in WT MEFs (Supplementary Information, Fig. 3f). However, neither T27N Arf6 (Fig. 4b, lower panel) nor Arf6 knockdown (data not shown) had any effect on GM1 endocytosis after detachment. Thus dominant-negative Arf6 and Arf6 depletion both indicate a requirement for Arf6 in raft exocytosis during replating.
The insensitivity of Cav1−/− cells to Arf6 inhibition supports the conclusion that membrane trafficking underlies the Arf6 requirement; however, Cav1 can inhibit signalling through its scaffolding domain or perhaps other mechanisms. We therefore compared WT Cav1 and phosphorylation-deficient Y14FCav1. This mutant contains an intact scaffolding domain and forms caveolae but does not support GM1 internalization in detached cells2. Transfection of Cav1−/− cells with WT Cav1 restored sensitivity to T27N Arf6, whereas Y14FCav1 was ineffective (Supplementary Information, Fig. 3a, b). Taken together, these data suggest that Arf6 controls the adhesion-regulated recycling of rafts from the recycling endosomes to the plasma membrane and identifies a new Arf6-dependent, Rab11- and Rab22-independent, pathway for the transport of lipid rafts out of recycling endosomes in spreading cells.
Activation and membrane targeting of Rac1
We next determined whether the Arf6-dependent transport of lipid rafts is necessary for the membrane targeting and activation of Rac1 during cell spreading28,29. As before, we compared WT and Cav1−/− cells. In WT MEFs, membrane targeting and activation of Rac1 were decreased in suspension (Fig. 4e, f). After 20 min on fibronectin, both the membrane targeting and activity of Rac1 were restored in cells expressing WT Arf6, whereas cells expressing T27N Arf6 showed a 50% decrease in membrane association of Rac1 and an 80% decrease in its activation. However, after 45 min the membrane targeting and activity of Rac1 were comparable in WT and T27N Arf6 cells (Fig. 4e, f). Arf6 knockdown also similarly affects Rac1 membrane levels in WT MEFs (Supplementary Information, Fig. 3g). No difference was observed in Rac1 activation and membrane association between non-transfected and WT Arf6-transfected cells (data not shown). Consistent with these results are the observations that cells eventually spread and that surface GM1 levels recover in cells expressing T27N Arf6 (Supplementary Information, Fig. 2e, f). In contrast to WT MEFs, membrane levels and activation of Rac1 remained high in detached Cav1−/− cells, did not change on readhesion, and were insensitive to T27N Arf6 (Fig. 4e, f). These results show that Arf6-dependent membrane raft trafficking is required for the adhesion-dependent regulation of Rac1.
Role of Arf6 activation
To further test the role of Arf6 in adhesion-dependent raft trafficking, we measured the effects of adhesion to fibronectin on Arf6 activation by using a pulldown assay30. Arf6 activity decreased by more than 60% after detachment and recovered rapidly on readhesion (Fig. 5a). Arf6 behaved in a qualitatively similar manner in Cav1−/− (Supplementary Information, Fig. 4a) and Chinese hamster ovary cells31.

(a) Top: WT MEFs that were stably adherent (SA), suspended for 90 min (susp.), or replated on fibronectin (10 μg ml−1) for 15 min (FN 15 min) or 30 min (FN 30 min) were lysed, and active Arf6 was pulled down on GST–GGA3 beads (GGA-PD). GGA-PD samples and the respective whole-cell lysates (WCL) were western blotted (WB) for Arf6. Numbers alongside blots represent molecular weight markers. Bottom: bands were quantified and normalized intensities were calculated relative to SA. Values are means and s.e.m. (n = 3). (b) MEFs expressing GFP-tagged WT Arf6 (WT) or HA-tagged T157A Arf6 (T157A) or non-transfected cells (control) were detached, surface-labelled with CTxB-Alexa 594 (GM1-CTxB) and held in suspension for 90 min. Images of cells with endocytosed GM1 were analysed to determine the intensity in the central recycling endosome pool (black columns) and in the rest of the cell (grey columns). Values are means and s.e.m. (n = 15; two independent experiments). (c) Top: suspended non-transfected (con.) and WT and T157A Arf6-expressing cells were replated on fibronectin (2 μg ml−1) for 15 min. Cells were surface-labelled with CTxB and bound CTxB was detected by western blotting (WB). Tubulin served as loading control. Samples were run on the same gel. Bottom: CTxB band intensity was quantified, normalized to tubulin and calculated relative to adherent control cells. Values are means and s.e.m., n = 3. (d) Cells expressing HA-tagged T157A Arf6 with endocytosed GM1-CTxB (as in b) were replated on fibronectin (2 μg ml−1) for 15 min, fixed and stained for HA. Arrows indicate areas of co-labelling suggestive of transport vesicles and ruffling edges. Uncropped images of the scans in a and c are shown in Supplementary Information, Fig. 7.
We next tested whether Arf6 activation is sufficient to drive GM1 from the recycling endosomes to the plasma membrane. Cells were transfected with WT Arf6 or an activated, fast-cycling mutant of Arf6 (T157A) that spontaneously exchanges nucleotide and shows lower toxicity than GTPase-deficient Q67L Arf6 (ref. 32). Neither T157A nor WT Arf6 significantly affected surface GM1 levels in adherent cells (Fig. 4c). The initial rate of GM1 endocytosis after detachment was also unaffected (Supplementary Information, Fig. 4e). However, after longer periods in suspension, endocytosed CTxB was markedly redistributed towards the cortical region in cells expressing T157A Arf6, in comparison with cells expressing WT Arf6 and control cells (Fig. 5b). These data suggest that active Arf6 drives the exit of CTxB from recycling endosomes.
Careful examination of CTxB-Alexa 594 images in cells expressing T157A Arf6 showed most of the cortical CTxB in vesicles under the plasma membrane rather than at the cell surface (Fig. 5b). To confirm this, cells expressing WT or active Arf6 were surface-labelled with CTxB, and the bound protein was quantified by western blotting (Fig. 5c). Little change in surface levels of GM1 was observed in adherent cells under any of these conditions. After 90 min in suspension, surface GM1 decreased in all of the cells, with T157A Arf6 causing only a roughly twofold increase in surface GM1 compared with control suspended cells, which was still about 2.6-fold less than in adherent cells. Cells expressing haemagglutinin (HA)-tagged T157A Arf6 were then prelabelled with CTxB, replated, fixed and stained for HA. CTxB strongly colocalized with T157A Arf6 in distinct vesicles, plasma membrane ruffles and lamellipodia (Fig. 5d; Pearson coefficient 0.69 ± 0.04). We also noticed that cells expressing T157A Arf6 spread faster than control cells (Supplementary Information, Fig. 4b) and had marginally higher Rac1 activation, although Rac1 membrane association was similar (Supplementary Information, Fig. 4c, d). Taken together, these results indicate that adhesion-dependent activation of Arf6 drives the exit of rafts from the recycling endosomes during cell spreading. However, additional adhesion-dependent steps may be needed for their efficient fusion with the plasma membrane.
Microtubules facilitate raft return to the plasma membrane
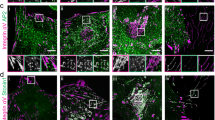
Because MTs mediate the movement of CTxB to the recycling endosomes and also target active lamellipodia33, we examined whether MTs might regulate the movement of raft components back to the plasma membrane during spreading. Cells were prelabelled with CTxB, replated on fibronectin and stained for β-tubulin. CTxB-containing vesicles lying between the recycling endosomes and the cell edge almost always colocalized with microtubules (Fig. 6a). To test this association functionally, cells were treated with nocodazole. WT and Cav1−/− MEFs were again compared, to assess the importance of lipid raft trafficking. We also took advantage of the fact that when cells were pretreated with nocodazole while adherent and were then detached, CTxB remained in the cortical region (Fig. 1a) with little localization to Rab11-positive recycling endosomes (Supplementary Information, Fig. 5a; Pearson coefficient 0.15 ± 0.05). In contrast, addition of nocodazole 90 min after detachment showed endocytosed CTxB in dispersed Rab11-positive structures (Supplementary Information, Fig. 5a; Pearson coefficient 0.50 ± 0.05). However, after both treatments GM1 was removed from the surface (Supplementary Information, Fig. 5b). When replated on fibronectin, spreading of WT cells was strongly inhibited by pretreatment with nocodazole while adherent but only moderately by the addition of nocodazole 90 min after detachment (Fig. 6b). By contrast, Cav1−/− cells were barely affected by either treatment. Treatment of WT MEFs with nocodazole after detachment also strongly decreased surface GM1 levels compared with nocodazole-pretreated or control cells (Fig. 6c). Addition of nocodazole after detachment also lowered Rac1 membrane targeting and activation, compared with untreated or nocodazole-pretreated cells (Supplementary Information, Fig. 5c, d). Cav1−/− cells were again resistant (Supplementary Information, Fig. 5c, d). The morphology of the nocodazole-treated cells was not entirely normal, but this result is consistent with MTs affecting the cytoskeleton through additional mechanisms such as focal adhesion turnover and regulation of Rho GTPases34,35. Nevertheless, these results show that regulation of exocytic trafficking of rafts in readherent cells is an important mechanism by which MTs regulate cell spreading.

(a) WT MEFs prelabelled with CTxB-Alexa 594 (GM1-CTxB) were held in suspension for 90 min and replated on fibronectin (2 μg ml−1) for 15 min. Fixed cells were stained for β-tubulin. Confocal images from multiple planes are shown as three-dimensional reconstructions. The right panel shows boxed regions of interest (1 and 2). (b) WT MEFs and Cav1−/−; MEFs were untreated (control) or treated with nocodazole (10 μM) before detachment (pre NOC) or after being suspended for 90 min (post NOC). Cells were replated on fibronectin (2 μg ml−1) for 15 min, fixed and stained for F-actin. Cell surface areas were determined from images. Values are means and s.e.m. for n = 25 cells, in arbitrary units; three independent experiments gave similar results. (c) Readherent cells were incubated with CTxB and bound CTxB was detected by western blotting (WB). Numbers alongside blot represent molecular weight markers. Samples were run on the same gel. Bands were quantified (bottom) and normalized for tubulin. Values are means and s.e.m. relative to WT MEF control cells (n = 3). Uncropped images of the scans in c are shown in Supplementary Information, Fig. 7.
Discussion
Previous studies showed that GM1-enriched lipid rafts were removed from the plasma membrane through caveolar endocytosis when anchorage-dependent cells were detached from the substratum2. Endocytosed rafts eventually coalesced in a distinct perinuclear region2. Integrin-mediated adhesion triggered their return to the plasma membrane. We now define this compartment as the recycling endosome and show that, in suspended cells, endocytosed rafts are transported there in a microtubule-dependent manner. Adhesion to fibronectin triggers rapid recycling of these membranes out of the recycling endosome by a new Arf6-dependent, Rab11- and Rab22-independent, pathway, also along microtubules. Interestingly, we observed little or no colocalization of rafts with caveolin in exocytic vesicles (Supplementary Fig. 6a), indicating that this pathway is also independent of caveolin.
However, Arf6 seemed insufficient for efficient delivery of raft components back to the plasma membrane; active Arf6 caused only a modest increase in their surface levels. Additional integrin-regulated step(s) may therefore be required for the docking and/or fusion of exocytic vesicles with the plasma membrane. This sequence of events is shown diagrammatically in Fig. 7.

Vesicles are transported along microtubules to recycling endosomes. Replating triggers the activation of Arf6 (Arf6*), which drives the movement of rafts from the recycling endosomes along the microtubules to the cortical region of the cell. Other, unknown, adhesion-dependent events complete the return of rafts to the plasma membrane.
In adherent cells under steady-state conditions, endocytosis of GM1-enriched rafts can occur through clathrin-dependent, caveolin-dependent or clathrin- and caveolin-independent pathways21,36,37. In adherent cells, raft markers internalized through caveolae are trafficked predominently to the Golgi17,38 and the SER20 and partly to the recycling endosome17. By contrast, cell detachment triggers the rapid and virtually complete internalization of raft components that are transported almost exclusively to the recycling endosome. This difference in post-endocytic routing probably reflects the rapid uptake of a large volume of raft components through a specific pathway activated by detachment. Similarly, although multiple pathways may deliver rafts from recycling endosomes to the plasma membrane at steady state, the present study identifies an Arf6-dependent, Rab11- and Rab22-independent, pathway as the primary exocytic pathway triggered by readhesion. Indeed, the restoration of surface GM1 and of Rac1 function and spreading at later times after plating, when Arf6 is inhibited, are consistent with the existence of additional, less efficient exocytic pathways.
Previous studies identified lipid rafts as membrane binding sites for active Rac1 and probably for related small GTPases4. Rafts are enriched in lamellipodia and are required for cell spreading1,4,39. Our data show that blocking the return of rafts to the plasma membrane by inhibiting Arf6 function or microtubule assembly inhibited the membrane targeting and activation of Rac1, and concomitantly impaired cell spreading in WT MEFs. Importantly, Cav1−/− cells or Y14FCav1-expressing cells, in which rafts are not internalized after detachment, are resistant to these inhibitors. Thus, transport of rafts to the leading edge emerges as a crucial mechanism by which both Arf6 and MTs control Rac-mediated cell spreading.
Microtubule inhibitors have highly variable effects on cell spreading, depending on the cell type and the exact protocol used14,40,41. Our studies show that Cav1 expression, its phosphorylation (Y14) and the timing of drug treatment could account for differences in effects of microtubule disruption on lipid raft trafficking and hence on cell spreading. Interestingly, both the disruption of MTs and the loss of Cav1 induce defects in polarity during migration12,33,42,43. It is therefore tempting to speculate that the microtubule-dependent transport of rafts to lamellipodia contributes to polarity during migration. Indeed, membrane binding sites for Rac1 within rafts localize to the lamellipodial edges1,4,5,8.
Arf6 was reported to regulate cell spreading by controlling the trafficking of Rac1 from a perinuclear compartment to the plasma membrane13,26. The nature and significance of this minor perinuclear pool of Rac1 were unclear, especially because abundant Rac1 is present in the cytoplasm bound to RhoGDI and can exchange rapidly with membrane-bound Rac1 (ref. 44). Our data suggest that it is not the delivery of Rac1 itself that Arf6 regulates, but rather the delivery of Rac1 membrane binding sites in lipid rafts. These data do not exclude additional modulation of Rac1 activity by Arf6 through the regulation of Rac GEFs45 or GTPase-activating proteins (GAPs) or the direct binding of Arf6 to Rac46. Arf6 could also affect Rac GEFs through effects on membrane targeting sites in lipid rafts47. Thus, trafficking of rafts seems to be a major regulatory pathway by which Arf6 controls Rac1 activation and cell spreading. These results differ from previous studies, in which the activation of Rac by growth factors did not require cell adhesion29, suggesting that different GEFs and GAPs have distinct requirements for raft membrane binding sites.
This study raises many new questions. Which motors drive centripetal movement of vesicles during endocytosis and centrifugal movement during exocytosis? How do raft and non-raft components coexist in the recycling endosome? How do membrane binding sites in lipid rafts regulate specific Rac GEFs and GAPs? Which Arf6 GEFs and effectors trigger the formation of raft-enriched exocytic vesicles from the recycling endosome during replating? How are they regulated by integrins and which additional integrin-regulated pathways control the final step of fusion with the plasma membrane? The answers to these questions are likely to illuminate a variety of integrin-regulated processes.
Methods
Reagents.
CTxB labelled with Alexa 594 (C22843) or Alexa 488 (C22841) were from Molecular Probes (Eugene, OR). Unlabelled cholera toxin B subunit (catalogue no. 227039), anti-cholera toxin antibody (catalogue no. 227040) and BFA (catalogue no. 203729) were obtained from Calbiochem (San Diego, CA). Latrunculin A (L5163), Nocodazole (M1404) and Alexa 488-transferrin were from Sigma (St Louis, MO, USA). Monoclonal anti-β-tubulin antibody (E7) was from the Developmental Studies Hybridoma Bank (Iowa City, IA). Anti-γ-tubulin and anti-caveolin-1 antibodies were from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-GM130 conjugated to fluorescein isothiocyanate (FITC) and anti-phospho-caveolin (Tyr 14) were purchased from Transduction Labs (San Jose, CA). Purified biotinylated aerolysin and anti-aerolysin antibody were from Protox Biotech (Victoria, Canada). Anti-Rac1 antibody was from Upstate Biotechnology (Lake Placid, NY). Anti-β1 integrin antibody was obtained from A. F. Horwitz (University of Virginia, Charlottesville, VA). Anti-AMF (autocrine motility factor) receptor antibody was obtained from I.R. Nabi (University of British Colombia, Vancouver, Canada). Anti-Arf1, anti-Arf3, anti-Arf4 and anti-Arf5 specific antibodies were obtained from R. Kahn (Emory University School of Medicine, Atlanta GA). Temperature-sensitive VSVG–GFP was obtained from J. Lippincott-Schwartz (NICHHD, NIH, Bethesda, MD). Protein kinase D1 kinase-dead mutant (PKD∼KD), PKD∼KD K618N and PKD∼KD P155/287G mutant constructs were obtained from V. Malhotra (University of California, San Diego, CA). WTRab11–GFP, (S25N) Rab11, WT Rab22a, dominant-negative (S19N) Rab22a, WT Cav1, Y14FCav1, WT Arf6, dominant-negative (T27N) Arf6, fast-cycling (T157A) Arf6 constructs and the monoclonal anti-Arf6 antibody were as described2,24,30,32,48. The RNA interference sequence 5′-AGCTGCACCGCATTATCAA-3′ of rat Arf6 mRNA (GenBank accession number NM_024152), used to generate complementary small interfering RNA (siRNA) oligonucleotides annealed and inserted into pSUPER.gfp/neo, were as described49.
Tissue culture.
Mouse embryonic fibroblasts from Cav1−/− and Cav1+/+ littermate mice (provided by R. Anderson, University of Texas Health Sciences Center, Dallas, TX, USA) were cultured in DMEM medium with 10% fetal bovine serum, penicillin and streptomycin (Invitrogen, Carlsbad, CA, USA). Cells (106) were transfected with 40 μg of DNA (5 μg of DNA plasmid plus 35 μg of salmon sperm DNA) by electroporation with the GenePulser Xcell (Bio-Rad Laboratories, Hercules, CA, USA). Cells were incubated for 6 h with 5 mM sodium butyrate to promote protein expression. If required, cells were serum-starved for 12 h after electroporation in medium with 0.2% fetal bovine serum. Cells analysed about 24 h after electroporation generally showed 90–95% transfection efficiency. Arf6 (60 μg) or scrambled dsRNA (60 μg) in pSUPER vector were electroporated into cells; cells were incubated for 72 h and then serum starved as above.
For experiments, cells were detached with 1 × trypsin, treated with soybean trypsin inhibitor, washed and then held in suspension with 1% methylcellulose before replating on fibronectin-coated coverslips. For aerolysin labelling experiments, cells were detached with trypsin-free Accutase (protease and collagenolytic activity; Innovative Cell Technologies Inc. (San Diego, CA)and processed as above. Fibronectin was used at 10 μg ml−1 except in time course experiments, in which the concentration was reduced to 2 μg ml−1 to facilitate the analysis of rates of exocytosis and spreading, as indicated in figure legends.
Analysis of surface CTxB and aerolysin.
Suspended or adherent cells were placed on ice for 15 min and then incubated with 10 μg ml−1 unlabelled CTxB (Calbiochem) or 1 μg ml−1 biotinylated aerolysin for 30 min. Cells were washed with cold PBS and 2 × 105 cells were lysed in 100 μl of SDS sample buffer. Cell-equivalent amounts of lysate were resolved by SDS–PAGE, transferred to nitrocellulose and blocked with 5% nonfat dried milk in TBS containing 0.5% Tween 20 (TBS-T). Blots were incubated with anti-CTxB antibody (1 μg ml−1) or anti-aerolysin antibody (0.5 μl ml−1) followed by anti-goat or anti-mouse antibody conjugated to horseradish peroxidase, respectively, and developed with the ECL+ enhanced chemiluminescence detection system (Amersham, Buckinghamshire, UK). Blots were stripped using Re-blot Plus stripping solution (Chemicon International, Temecula, CA) and re-probed with monoclonal anti-β-tubulin antibody as a loading control. Bands were quantified by densitometry with Image J software (NIH). CTxB band intensities were normalized to tubulin.
Localization of CTxB.
Stably adherent cells were placed on ice for 15 min then incubated for 15 min with 10 μg ml−1 CTxB-Alexa 594 or CTxB-Alexa 488, as indicated, in PBS. Cells were detached, held in suspension and replated as above, and the movement of endocytosed CTxB was studied. In suspended or replated cells, surface GM1 was detected by incubating cells for 15 min on ice with 10 μg ml−1 CTxB-Alexa 594 in PBS before fixation in 3.5% paraformaldehyde. Labelled cells were permeabilized for 15 min in PBS containing 3% BSA and 0.05% Triton X-100, then blocked for 1 h with 3% BSA in PBS. Cells were then stained with anti-AMF-R antibody, followed by anti-rat Alexa 488 antibody (Molecular Probes), with the anti-γ-tubulin antibody, followed by anti-mouse Alexa 488 antibody, or directly with FITC-conjugated anti-GM130 antibody. Cells expressing HA-tagged Arf6 were incubated with anti-HA antibody, followed by anti-mouse IgG-Alexa 488. For double labelling of CTxB-Alexa 568 with tubulin, cells were fixed for 10 min with cold 80% methanol at −20 °C, stained with anti-β-tubulin monoclonal antibody and anti-mouse IgG Alexa 488 secondary antibody. Cells were mounted in Fluoromount-G (Southern Biotech, Birmingham, AL[), observed with a Zeiss LSM 510 laser confocal microscope with a 40× or 60× objective and analysed with either a Zeiss LSM Image Browser or Image J software.
Quantification of intracellular CTxB localization.
Cells were surface-labelled with Alexa-CTxB, suspended, fixed and mounted as detailed above. Confocal images were recorded, being careful to avoid pixel saturation. With the use of Image J software, two thresholds were set, one to map the entire cell and the other to map the compact internal pool of endocytosed CTxB. The tracing tool was then used to select edges of the thresholded areas. Total intensity within the thresholded areas was determined, and the percentage of CTxB within the internal pool and the rest of the cell was calculated.
Co-labelling of cells with transferrin and CTxB.
Stably adherent cells were incubated for 30 min with 20 μg ml−1 transferrin-Alexa 594 (Sigma), washed, and chilled on ice for 15 min. Cells were then incubated with 10 μg ml−1 Alexa 488-CTxB (Molecular Probes) in DMEM for 30 min. Cells were washed with DMEM, detached and held in suspension with 1% methylcellulose in DMEM. At the end of the incubation they were fixed or replated on fibronectin as described above.
Quantification of endocytosed transferrin distribution.
Cells labelled with transferrin-Alexa 594 as above and replated were fixed and mounted; confocal images were recorded, being careful to avoid pixel saturation. With the use of Image J software, two thresholds were set, one to map the entire cell and the other to map the endocytosed transferrin; the thresholded area for each was calculated. The area occupied by transferrin was calculated as a percentage of the total cell area.
Drug treatments.
Stably adherent cells were incubated for 30 min with 1 μM latrunculin A, detached with trypsin, held in suspension for 90 min with 1% methylcellulose, fixed or replated on fibronectin and fixed with 3.5% paraformaldehyde. For treatment with BFA, cells suspended for 90 min were incubated for 30 min with 1 μg ml−1 BFA in suspension and fixed immediately or replated on fibronectin before fixation. For early nocodazole treatment, stably adherent cells were incubated for 10 min with 10 μM nocodazole, detached with trypsin, washed with DMEM, and held in suspension for 90 min with 1% methylcellulose in DMEM in the continuous presence of the drug. For late nocodazole treatment, 10 μM nocodazole was added to cells after 90 min in suspension. Control cells were left untreated. All cells were incubated for a further 30 min in suspension, washed to remove methylcellulose and replated on coverslips coated with 2 μg ml−1 fibronectin with or without nocodazole. After 15 min, cells were washed, fixed and mounted as before. Cells were observed with a Nikon Diaphot TMD fluorescence microscope using a 60× oil-immersion objective and photographed with a Coolsnap HQ camera (Roper Scientific, Tucson, AZ).
Arf6 activity assay.
Arf6 activity was measured as described previously30. Stably adherent cells were detached, held in suspension for 2 h and replated on 10 μg ml−1 fibronectin. Cells were lysed on ice in buffer containing 50 mM Tris-HCl pH 7.5, 100 mM NaCl, 2 mM MgCl2, 1% Triton X-100 and protease and phosphatase inhibitor cocktails (Calbiochem). Lysates were centrifuged for 30 min at 13,000g and incubated for 30 min with 40 μg GST–GGA3 (GST-fused Arf binding domain of GGA3)plus 10 μl of glutathione–Sepharose beads (Amersham) at 4 °C. Beads were washed three times in lysis buffer and eluted with SDS sample buffer. Arf6 was detected by western blotting with anti-Arf6 antibody30.
Subcellular fractionation.
Cells were washed with cold PBS, scraped, and homogenized in 500 μl of buffer containing 10 mM Tris-HCl pH 7.5, 5 mM MgCl2, 1 mM dithiothreitol, 0.25 M sucrose and a cocktail of protease inhibitors (Sigma). Nuclei and unbroken cells were removed by centrifugation for 10 min at 1000g and 4 °C, and the postnuclear supernatant was centrifuged for 1 h at 100,000g and 4 °C to sediment the membranes; 60% of the membrane fraction and 1.6% of the cytosolic fraction were analysed by western blotting for Rac1 and β1 integrin. Rac1 levels in the membrane fraction were normalized to β1 integrin levels.
Rac1 activity assay.
Rac1 pulldown assays for GTP loading were performed essentially as described29; 60% of GST–p21-binding domain (PBD)-bound active Rac1 and 1.6% of whole-cell lysates were analysed by western blotting and bands were quantified by densitometry. The amount of active Rac was calculated relative to total Rac for each sample.
Measuring cell spreading.
Images of adherent cells were analysed with Image J software. Thresholds were set to map the entire cell, the tracing tool was used to select the edge of each thresholded cell. The total area within the mapped edge for each cell was determined.
Quantification of early CTxB endocytosis in suspended cells.
Cells photographed with the laser confocal microscope were analysed with Image J. By thresholding and using the 'percentage shrink' macro, 20% of the cell area from the cell periphery (about 0.5–0.7 μm) was defined as the cell edge. The central 40% area of the cell was defined as the cell centre. Intensities in these regions were measured, normalized to area and are presented as percentages of total CTxB intensity.
Statistical significance.
Comparisons between data points were done with Student's t-test (Sigmaplot Statistical Analysis Software).
Fluorescent image colocalization analysis.
Images of cells photographed with the laser confocal microscope at both wavelengths were analysed with Image J software, and the Pearson coefficient was determined with the colocalization threshold plugin. Intensity quantification along a defined line for each wavelength was determined with the 'plot profile' function. Lists of values obtained were plotted in sigmaplot.
Note: Supplementary Information is available on the Nature Cell Biology website.
References
del Pozo, M. A. et al. Integrins regulate Rac targeting by internalization of membrane domains. Science 303, 839–842 (2004).
del Pozo, M. A. et al. Phospho-caveolin-1 mediates integrin-regulated membrane domain internalization. Nature Cell Biol. 7, 901–908 (2005).
Palazzo, A. F., Eng, C. H., Schlaepfer, D. D., Marcantonio, E. E. & Gundersen, G. G. Localized stabilization of microtubules by integrin- and FAK-facilitated Rho signaling. Science 303, 836–839 (2004).
Vasanji, A., Ghosh, P. K., Graham, L. M., Eppell, S. J. & Fox, P. L. Polarization of plasma membrane microviscosity during endothelial cell migration. Dev. Cell 6, 29–41 (2004).
Golub, T. & Pico, C. Spatial control of actin-based motility through plasmalemmal PtdIns(4,5)P2-rich raft assemblies. Biochem. Soc. Symp. 72, 119–127 (2005).
Simons, K. & Toomre, D. Lipid rafts and signal transduction. Nature Rev. Mol. Cell Biol. 1, 31–39 (2000).
Hancock, J. F. Lipid rafts: contentious only from simplistic standpoints. Nature Rev. Mol. Cell Biol. 7, 456–462 (2006).
Manes, S. et al. Membrane raft microdomains mediate front–rear polarity in migrating cells. EMBO J. 18, 6211–6220 (1999).
Gaus, K., Le Lay, S., Balasubramanian, N. & Schwartz, M. A. Integrin-mediated adhesion regulates membrane order. J. Cell Biol. 174, 725–734 (2006).
Donaldson, J. G. Multiple roles for Arf6: sorting, structuring, and signaling at the plasma membrane. J. Biol. Chem. 278, 41573–41576 (2003).
D'Souza-Schorey, C. & Chavrier, P. ARF proteins: roles in membrane traffic and beyond. Nature Rev. Mol. Cell Biol. 7, 347–358 (2006).
Sabe, H. Requirement for Arf6 in cell adhesion, migration, and cancer cell invasion. J. Biochem. (Tokyo) 134, 485–489 (2003).
Radhakrishna, H., Al-Awar, O., Khachikian, Z. & Donaldson, J. G. ARF6 requirement for Rac ruffling suggests a role for membrane trafficking in cortical actin rearrangements. J. Cell Sci. 112, 855–866 (1999).
Cheung, H. T. & Terry, D. S. Effects of nocodazole, a new synthetic microtubule inhibitor, on movement and spreading of mouse peritoneal macrophages. Cell Biol. Int. Rep. 4, 1125–1129 (1980).
Gundersen, G. G. & Bulinski, J. C. Selective stabilization of microtubules oriented toward the direction of cell migration. Proc. Natl Acad. Sci. USA 85, 5946–5950 (1988).
Krendel, M., Zenke, F. T. & Bokoch, G. M. Nucleotide exchange factor GEF-H1 mediates cross-talk between microtubules and the actin cytoskeleton. Nature Cell Biol. 4, 294–301 (2002).
Le, P. U. & Nabi, I. R. Distinct caveolae-mediated endocytic pathways target the Golgi apparatus and the endoplasmic reticulum. J. Cell Sci. 116, 1059–1071 (2003).
Prigozhina, N. L. & Waterman-Storer, C. M. Protein kinase D-mediated anterograde membrane trafficking is required for fibroblast motility. Curr. Biol. 14, 88–98 (2004).
Presley, J. F. et al. ER-to-Golgi transport visualized in living cells. Nature 389, 81–85 (1997).
Pelkmans, L., Kartenbeck, J. & Helenius, A. Caveolar endocytosis of simian virus 40 reveals a new two-step vesicular-transport pathway to the ER. Nature Cell Biol. 3, 473–483 (2001).
Sabharanjak, S., Sharma, P., Parton, R. G. & Mayor, S. GPI-anchored proteins are delivered to recycling endosomes via a distinct cdc42-regulated, clathrin-independent pinocytic pathway. Dev. Cell 2, 411–423 (2002).
Ullrich, O., Reinsch, S., Urbe, S., Zerial, M. & Parton, R. G. Rab11 regulates recycling through the pericentriolar recycling endosome. J. Cell Biol. 135, 913–924 (1996).
Yamashiro, D. J. & Maxfield, F. R. Acidification of endocytic compartments and the intracellular pathways of ligands and receptors. J. Cell Biochem. 26, 231–246 (1984).
Weigert, R., Yeung, A. C., Li, J. & Donaldson, J. G. Rab22a regulates the recycling of membrane proteins internalized independently of clathrin. Mol. Biol. Cell 15, 3758–3770 (2004).
D'Souza-Schorey, C. & Chavrier, P. ARF6 targets recycling vesicles to the plasma membrane: insights from an ultrastructural investigation. J. Cell Biol. 140, 603–616 (1998).
Zhang, Q., Calafat, J., Janssen, H. & Greenberg, S. ARF6 is required for growth factor- and rac-mediated membrane ruffling in macrophages at a stage distal to rac membrane targeting. Mol. Cell. Biol. 19, 8158–8168 (1999).
Song, J., Khachikian, Z., Radhakrishna, H. & Donaldson, J. G. Localization of endogenous ARF6 to sites of cortical actin rearrangement and involvement of ARF6 in cell spreading. J. Cell Sci. 111, 2257–2267 (1998).
Price, L. S., Leng, J., Schwartz, M. A. & Bokoch, G. M. Activation of Rac and Cdc42 by integrins mediates cell spreading. Mol. Biol. Cell 9, 1863–1871 (1998).
del Pozo, M. A., Price, L. S., Alderson, N. B., Ren, X. D. & Schwartz, M. A. Adhesion to the extracellular matrix regulates the coupling of the small GTPase Rac to its effector PAK. EMBO J. 19, 2008–2014 (2000).
Santy, L. C. & Casanova, J. E. Activation of ARF6 by ARNO stimulates epithelial cell migration through downstream activation of both Rac1 and phospholipase D. J. Cell Biol. 154, 599–610 (2001).
Goldfinger, L. E., Ptak, C., Jeffery, E. D., Shabanowitz, J., Hunt, D. F., Ginsberg, M. H. RLIP76 (RalBP1) is an R-Ras effector that mediates adhesion-dependent Rac activation and cell migration. J. Cell Biol. 174, 877–888 (2006).
Santy, L. C. Characterization of a fast cycling ADP-ribosylation factor 6 mutant. J. Biol. Chem. 277, 40185–40188 (2002).
Schmoranzer, J., Kreitzer, G. & Simon, S. M. Migrating fibroblasts perform polarized, microtubule-dependent exocytosis towards the leading edge. J. Cell Sci. 116, 4513–4519 (2003).
Ren, Y., Li, R., Zheng, Y. & Busch, H. Cloning and characterization of GEF-H1, a microtubule-associated guanine nucleotide exchange factor for Rac and Rho GTPases. J. Biol. Chem. 273, 34954–34960 (1998).
Kawasaki, Y. et al. Asef, a link between the tumor suppressor APC and G-protein signaling. Science 289, 1194–1197 (2000).
Torgersen, M. L., Skretting, G., van Deurs, B. & Sandvig, K. Internalization of cholera toxin by different endocytic mechanisms. J. Cell Sci. 114, 3737–3747 (2001).
Nabi, I. R. & Le, P. U. Caveolae/raft-dependent endocytosis. J. Cell Biol. 161, 673–677 (2003).
Nichols, B. J. A distinct class of endosome mediates clathrin-independent endocytosis to the Golgi complex. Nature Cell Biol. 4, 374–378 (2002).
Caroni, P. New EMBO members' review: actin cytoskeleton regulation through modulation of PI(4,5)P2 rafts. EMBO J. 20, 4332–4336 (2001).
Pletjushkina, O. J. et al. Induction of cortical oscillations in spreading cells by depolymerization of microtubules. Cell Motil. Cytoskel. 48, 235–244 (2001).
Rosania, G. R. & Swanson, J. A. Microtubules can modulate pseudopod activity from a distance inside macrophages. Cell Motil. Cytoskel. 34, 230–245 (1996).
Grande-Garcia, A. et al. Caveolin-1 regulates cell polarization and directional migration through Src kinase and Rho GTPases. J. Cell Biol. 177, 683–694 (2007).
Etienne-Manneville, S. Actin and microtubules in cell motility: which one is in control? Traffic 5, 470–477 (2004).
Moissoglu, K., Slepchenko, B. M., Meller, N., Horwitz, A. F. & Schwartz, M. A. In vivo dynamics of Rac–membrane interactions. Mol. Biol. Cell 17, 2770–2779 (2006).
Santy, L. C., Ravichandran, K. S. & Casanova, J. E. The DOCK180/Elmo complex couples ARNO-mediated Arf6 activation to the downstream activation of Rac1. Curr. Biol. 15, 1749–1754 (2005).
Cotton, M. et al. Endogenous ARF6 interacts with Rac1 upon angiotensin II stimulation to regulate membrane ruffling and cell migration. Mol. Biol. Cell 18, 501–511 (2007).
Prag, S. et al. Activated ezrin promotes cell migration through recruitment of the GEF Dbl to lipid rafts and preferential downstream activation of Cdc42. Mol. Biol. Cell 18, 2935–2948 (2007).
Ren, M. et al. Hydrolysis of GTP on rab11 is required for the direct delivery of transferrin from the pericentriolar recycling compartment to the cell surface but not from sorting endosomes. Proc. Natl Acad. Sci. USA 95, 6187–6192 (1998).
Choi, S. et al. ARF6 and EFA6A regulate the development and maintenance of dendritic spines. J. Neurosci. 26, 4811–4819 (2006).
Acknowledgements
We thank Robert Nabi (University of British Colombia, Vancouver, Canada) for generously providing anti-AMF antibody; Richard Kahn (Emory University School of Medicine, Atlanta GA) for antibodies to Arfs 1, 3, 4 and 5; Jennifer Lippincott-Schwartz (NICHHD, NIH, Bethesda, MD) for VSVG-GEP; and Vivek Malhotra (University of California, San Diego, CA) for PKD constructs. Anti-tubulin antibody was obtained from the Developmental Studies Hybridoma Bank. This work was supported by a US Public Health Service grant RO1 GM47214 to M.A.S.
Author information
Authors and Affiliations
Contributions
N.B. designed and carried out experiments, analysed data and wrote the manuscript. D.W.S. carried out experiments. J.D.C. and J.E.C. provided reagents and advice. M.A.S. designed experiments, analysed data and wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary figures S1, S2, S3, S4, S5, S6 and S7 (PDF 3618 kb)
Rights and permissions
About this article
Cite this article
Balasubramanian, N., Scott, D., Castle, J. et al. Arf6 and microtubules in adhesion-dependent trafficking of lipid rafts. Nat Cell Biol 9, 1381–1391 (2007). https://doi.org/10.1038/ncb1657
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ncb1657
This article is cited by
-
CNK2 promotes cancer cell motility by mediating ARF6 activation downstream of AXL signalling
Nature Communications (2023)
-
A mobile endocytic network connects clathrin-independent receptor endocytosis to recycling and promotes T cell activation
Nature Communications (2018)
-
A cell-based high-throughput screening assay system for inhibitor compounds of antigen presentation by HLA class II molecule
Scientific Reports (2017)
-
Integrin-Dependent Regulation of Small GTPases: Role in Cell Migration
Journal of the Indian Institute of Science (2017)
-
Basigin-mediated redistribution of CD98 promotes cell spreading and tumorigenicity in hepatocellular carcinoma
Journal of Experimental & Clinical Cancer Research (2015)
