
Abstract
The Morris water maze (MWM) is a test of spatial learning for rodents that relies on distal cues to navigate from start locations around the perimeter of an open swimming arena to locate a submerged escape platform. Spatial learning is assessed across repeated trials and reference memory is determined by preference for the platform area when the platform is absent. Reversal and shift trials enhance the detection of spatial impairments. Trial-dependent, latent and discrimination learning can be assessed using modifications of the basic protocol. Search-to-platform area determines the degree of reliance on spatial versus non-spatial strategies. Cued trials determine whether performance factors that are unrelated to place learning are present. Escape from water is relatively immune from activity or body mass differences, making it ideal for many experimental models. The MWM has proven to be a robust and reliable test that is strongly correlated with hippocampal synaptic plasticity and NMDA receptor function. We present protocols for performing variants of the MWM test, from which results can be obtained from individual animals in as few as 6 days.
Similar content being viewed by others
Introduction
Many water mazes have been developed, but the one that is referred to as 'the water maze' was developed by Richard Morris1. The maze was designed as a method to assess spatial or place learning and herein will be referred to as the Morris water maze (MWM). Morris described the basic procedures in 1984 (ref. 2) and subsequently added details and procedures for assessing related forms of learning and memory3. Several characteristics have contributed to the prevalent use of the MWM. These include the lack of required pretraining, its high reliability across a wide range of tank configurations and testing procedures, its cross-species utility (rats, mice and humans (in a virtual maze4)), extensive evidence of its validity as a measure of hippocampally dependent spatial navigation and reference memory5, its specificity as a measure of place learning, and its relative immunity to motivational differences across a range of experimental treatment effects that are secondary to the central purpose of the task (genetic, pharmacological, nutritional, toxicological and lesion). Although the latter is a general characteristic shared by all water mazes6, the MWM capitalizes on this strength. For example, hippocampal and septohippocampal lesions in rats reliably induce hyperactivity, but such animals show deficits in the MWM7. At the opposite pole, treatments that induce hypoactivity can be dissociated from learning deficits in the MWM. For example, it has been shown that MWM learning impairments are independent of locomotor effects because land-based locomotor reductions did not affect swimming speed. Moreover when the experimental animals have deficits during probe trials, this further dissociates learning from performance because measures recorded on probe trials are insensitive to swimming speed8.
The use of the MWM in assessing learning and memory has been reviewed9,10, as has the relationship between performance in the MWM and both neurotransmitter systems and drug effects11. MWM performance has been linked to long-term potentiation (LTP) and NMDA receptor function12,13,14,15, making it a key technique in the investigation of hippocampal circuitry. In addition, it has been shown that there is involvement of the entorhinal and perirhinal cortices, as well as involvement of the prefrontal cortex, the cingulate cortex, the neostriatum, and perhaps even the cerebellum in a more limited way10.
Despite extensive use of the MWM, the task has not always been used optimally. Some of this stems from an under-appreciation for the aspects of the apparatus and testing procedures that are most salient for obtaining the best possible data. Here, we provide a description of the apparatus, its key features, and protocols that are effective and reliable for detecting drug/lesion-induced changes in spatial learning and memory16,17,18,19,20 or changes that arise as a result of genetic manipulations21,22,23,24,25,26,27,28. We also provide variations to the basic protocol that can be used to enhance assessment of spatial navigation and/or test for related types of learning (latent, discrimination, and cued learning or working memory).
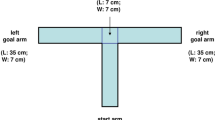
The MWM is not a maze in the usual sense — that is, it is not a labyrinth; rather, it is an open circular pool that is filled approximately half-way with water. The interior is made such that it is as close to being featureless as possible. It is a 'maze' in the sense that the animal must search in order to locate a relatively small goal (a hidden platform) that is submerged below the water surface and placed in a fixed location. The platform is camouflaged either by placing opacifying materials in the water (typically, tempera paint or polypropylene pellets), by creating a nearly invisible platform-to-background color match, or by using transparent platforms against a colored background, thereby making it indistinct given the low visual aspect ratio to the water as seen by the animal when swimming.
It is standard to designate two principal axes of the maze, each line bisecting the maze perpendicular to one another to create an imaginary '+'. The end of each line demarcates four cardinal points: North (N), South (S), East (E) and West (W). These are not true magnetic compass directions but refer to S being the experimenter's position, N being at the opposite point, E being to the experimenter's right and W being to the experimenter's left. Dividing the maze this way creates four equal quadrants. The platform is positioned in the middle of one of the quadrants. One can either keep the platform in one quadrant for all trials or test one-quarter of the animals with the platform in each of the quadrants. The latter approach counterbalances for possible quadrant effects. One can even use eight different platform positions22. The platform is usually located half-way between the center and the wall, regardless of the quadrant selected, although other arrangements are sometimes used29.
Test protocols
Spatial acquisition. Place or spatial learning is the most basic MWM procedure. The concept behind it is that the animal must learn to use distal cues to navigate a direct path to the hidden platform when started from different, random locations around the perimeter of the tank. If there are no proximal cues available, the use of distal cues provides the most effective strategy to accomplish this. Most protocols use four start locations: N, S, E and W. Animals are given a series of daily trials using a random or semi-random set of start locations. Semi-random start position sets are most common, such that the four positions are used, with the restriction that one trial each day is from each of the four positions. A few investigators use eight start locations30. One concern about the cardinal start positions is that they are not equidistant from the goal, creating short and long paths to the goal. Even in a large maze, a rat starting at E, with the goal located at SE, has a short path to the goal. There is no perfect solution to this problem. A partial solution that we have used is to use only distal start locations18. By this, we mean that if the goal is SE, then one can use start locations of N, W, NE and SW. Although not equidistant from the goal, these start positions are closer to being equal in length than using start positions that are adjacent to the goal. Another approach might be to use only two start positions, such as N and W only, but one must then be concerned that animals might memorize specific routes rather than use distal cues. A third approach is to use three start positions, each in quadrants other than the one containing the platform31, however only two of these are equal in length.
Table 1 illustrates a set of semi-randomly selected distal start positions for basic acquisition training, with the platform being located in the SW quadrant. These are designed so that the animal is not able to learn a specific order of right or left turns to locate the platform, while using each of the four start positions once each day. As can be seen in Table 1, the learning trials are conducted over 5 days, with 4 trials per day. The interval between trials can vary from 10–15 s to 5–15 min. If an animal fails to find the platform within the allotted time, it is usually picked up and placed on the platform for ∼15 s, although some prefer to guide the animal to the goal based on evidence that it is the middle portion of the swim path that seems to be most important in learning how to navigate to the goal32.
To assess reference memory at the end of learning, a probe (transfer) trial is given. The most common method is to administer one probe trial 24 h after the last acquisition day. With some procedures, the probe trial is administered immediately following the last learning trial; however, this cannot differentiate between short- and long-term memory, as it may reflect memory for the most recent training session. A long interval between the last training trial and the probe trial is essential if reference memory is to be determined independent of the memory of the last training session.
Additional probe trials are sometimes interspersed during the learning phase: these are often given before the first learning trial of the day. These additional probe trials may help to determine the rate of memory consolidation, as this allows the gradual emergence of goal quadrant preference to be seen across days. However, caution should be exercised not to conduct too many probe trials as these are extinction trials and may slow the rate of learning.
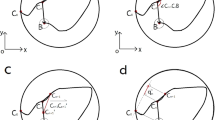
Spatial reversal. It is increasingly common and frequently informative to relocate the platform to another quadrant (usually the opposite one) and administer another set of four trials per day for 5 additional days (Table 1). This is often called reversal learning, although the term is not precise, as swimming to an opposite quadrant is not the mirror image of the initial problem as it is in a T-maze. Reversal learning in the MWM reveals whether or not animals can extinguish their initial learning of the platform's position and acquire a direct path to the new goal position. Tracking patterns typically reveal that mice swim to the previous location first, then begin to search in an arching pattern to reach the new goal (Fig. 1). Even after multiple trials, mice do not entirely abandon their initial learning strategy and begin trials by starting to move towards the original platform position, then turn and swim more directly to the new goal. Rats, on the other hand, rapidly switch their search strategies to the new goal position (Fig. 2). In fact, rats switch away from the old goal location so rapidly that return visits to the original platform location above chance (i.e., 25%) cannot be seen in the average of the first four trials on reversal day 1 but may be seen on individual trials within the first day of reversal testing. As in the acquisition phase, at the end of the reversal phase, a reversal probe trial is given 24 h later.

The results were averaged across four trials per day (mean ± s.e.m.) in untreated C57BL male mice during the reversal phase of learning — that is, after 6 days of acquisition training in which the platform was in the SW quadrant. During the trials shown (a–d), the platform was moved from the SW to the NE quadrant. The gradual reduction in percent time can be seen in the 'old' or previous SW goal quadrant and the gradual shift to the 'new' NW goal position (tank diameter, 122 cm).

The results were averaged across four trials per day (mean ± SEM) in untreated Sprague–Dawley male rats during the reversal phase of learning, as in Figure 1. The platform was in the SW quadrant during acquisition training and, during the trials shown (a–d), the platform was moved from the SW to the NE quadrant. As can be seen, rats show a rapid switch from the 'old' SW position to the 'new' NE platform position without perseverating on the original platform quadrant (tank diameter, 210 cm).
Spatial double-reversal with a smaller platform. Many variations can be added to the basic MWM procedures and these can add valuable information for understanding the deficits that are observed or may even unmask more subtle deficits that are not seen during acquisition or reversal learning. One procedure that has been effective in our hands has been to move the platform again, either back to the original goal (double-reversal) or to a different quadrant (shift), but with an additional change: use of a smaller platform17. For example, if the starting platform is 10 × 10 cm, the reduced platform may be 5 × 5 cm. This reduction in platform size taxes the spatial accuracy requirements of the animal and has revealed the effects of some drugs or doses that are not seen during acquisition or reversal16,20. A reduced platform probe trial is also given 24 h after the end of this phase of testing.
Repeated learning. Another procedure is to conduct a set of reversal or shift phases serially19. This allows an examination of the animals' flexibility in their ability to learn across multiple phases of new learning. The data also demonstrate the effects of moving the platform to different quadrants. For example, if the platform is shifted to an adjacent quadrant, new learning is more rapid than if shifted to an opposite quadrant19.
Spatial working memory. The procedures described above are for the assessment of trial-independent learning (that is, the goal does not move from trial to trial during a given phase of testing). To assess working or trial-dependent learning and memory, a different method is required. In this procedure, which is also called matching-to-sample, the platform is relocated every day and the animal is given two trials (or more) per day (see Table 2). On each day, the first trial represents a sample trial. During the sample trial, the animal must learn the new location of the platform by trial-and-error. Trial 2 (or any successive trial) is the test or matching trial in which savings in recall between Trial 1 and Trial 2 are measured. Trial 2 begins after a 15-s inter-trial interval. If the animal recalls the sample trial, it will swim a shorter path to the goal on the second trial. As the platform is moved daily, no learning of platform position from the previous day can be transferred to the next day's problem; hence, recall on each day during Trial 2 is dependent on that day's sample trial and measures only temporary or working memory.
Discrimination learning. The MWM can also be used to assess visual discrimination learning3. In this procedure, two visible platforms are used that are distinct from one another such as one being white and one being black. One is the standard fixed platform that is raised above the water and the other platform floats from a tether. The task for the subject is to learn which platform can be used for escape from the water and which cannot. The accuracy of the animal's choice across successive trials is an index of its ability to differentiate the stimulus information of the 'true' goal relative to the 'false' goal.
Latent learning. In latent learning, the idea is to place the animal on the platform before each trial rather than after. This will allow one to determine how much of the spatial learning stems from navigating to the platform compared with orientation to the goal once there. Morris has described this procedure elsewhere3.
Cued learning. A control condition that is frequently used in the MWM is to test the animals for their ability to learn to swim to a cued goal. In this procedure, curtains are closed around the maze to reduce the availability of distal cues. The curtains interfere with the animal's access to distal cues that could be used to spatially navigate. The platform is the same as in the hidden platform version, except that it is either elevated above the water surface19 or is kept submerged but a 'flag' is mounted that extends above the water surface by approximately 12 cm (ref. 33). Although both methods work, we find that the version with a flag seems to be more efficient, as it is readily recognizable from across the pool, whereas the protruding platform may not be. This 'cue' is designed to allow the animal a direct line-of-sight to the platform's location. To ensure that the animal is using this proximal cue to locate the platform, the location of the goal and the start are both moved to new positions during each trial (Table 3). In this way, the subject cannot use distal cues to solve the problem. The only cue that reliably indicates the location of the platform relative to the start is the cue that is attached to the platform. Morris introduced this as a control procedure as part of his original description of the test1. Unfortunately, this procedure is all too often omitted, yet its value is unmistakable. If subjects are impaired in cued learning, there is a potentially serious concern about whether a spatial deficit is present. This is because cued learning requires the same basic abilities (intact eyesight, motoric ability (swimming), basic strategies (learning to swim away from the wall, learning to climb on the platform)) and the same motivation (escape from water) as the spatial version of the task. Therefore, if the subject cannot perform the cued task, doubt is cast on its capacity to learn using distal cues in the spatial version. This task can be administered before or after the spatial version, but administering it before has advantages, especially for mice. Some animals find the platform, but then jump back into the water and continue searching. Presumably, this reflects an effort to find another route of escape. In the first few trials, some animals are sufficiently activated by being in the water that it is not always clear that they recognize that the platform is an escape when they first locate it. Therefore, testing animals first in cued trials eliminates the problem of animals not acquiring the appropriate subordinate skills before they are presented with the spatial version of the task.
Control procedures
Cued learning is basically a control procedure, but it is not the only one available. For example, Cain34,35,36 has shown that some drugs interfere with sensorimotor function and this can interfere with the animal's ability to recognize that the platform is the goal. He has proposed several ways of determining whether sensorimotor interference is occurring. He suggests measuring thigmotaxis, or the tendency to cling or follow the wall around the outer perimeter of the tank, as one index to reflect that the animal is not problem-solving. Excessive thigmotaxis (especially in rats) indicates that the animal is not focusing on the task appropriately because one of the first things that animals have to learn is that there is no escape located around the perimeter of the tank. Having learned this, most animals swim away from the wall and then, by weaving or looping search patterns, eventually find the platform. Not learning this basic approach indicates that the animal may not have adequate awareness of its surroundings. Other measures of impaired sensorimotor interference are excessive jump-offs, swimovers and/or deflections. Rats that reach the platform but do not climb on it, or do not stay on it, are not acquiring the requisite association between the platform and escape. Some of these behaviors may be seen during early trials even in control animals, but these usually disappear within a day. If such behaviors are more frequent in the experimental group, however, questions should be raised about whether spatial learning can be satisfactorily assessed.
Cain has suggested that one way to solve the problem of sensorimotor interference is to compare two groups of experimental animals: one tested in the standard procedure and one pre-trained using a 'non-spatial' training procedure. The pre-trained group is given a series of hidden platform trials in which the start and goal are moved randomly on every trial, as is done in cued learning, but here the curtains are left open and the platform is hidden. The task cannot be solved using spatial navigation because of the randomized start-goal combinations but it teaches the subject the basic task requirements — namely, the escape can only be found by searching, the goal is located somewhere away from the wall and the platform is the goal. Cain34,35,36,37,38,39, Morris15 and Whishaw40 have demonstrated that 'non-spatial' pretraining can separate components of learning that are not spatial from those that are and this, in turn, can change the interpretation of the findings. As non-spatial or strategy pretraining and cued training both have the effect of teaching animals the basic task requirements and tend to eliminate behaviors such as swimovers, jump-offs and even diving, it can be helpful to conduct cued trials first. Strategy pre-training is not usually necessary unless the data show that thigmotaxis or platform recognition behaviors indicate that sensorimotor problems are present. Acute pharmacological studies, however, may need non-spatial pretraining to ensure that non-cognitive effects are not interfering with maze performance, whereas delayed or long-term drug effect studies may not need this extra procedure.
Another approach to determine whether or not animals have any underlying sensorimotor deficits is to assess swimming speed. Often this is done in the maze during learning trials41. Alternatively, one can pre-test animals in a separate apparatus, such as a long, straight swimming channel. We use a 15 × 244 cm water-filled channel with an escape ladder or platform at one end33, although shorter channels have also been used31. This task requires virtually no searching, hence virtually no learning. The first one or two trials serve to acclimate them to swimming and the rats quickly recognize that the escape can be found by simply swimming from one end to the other. During subsequent trials (a total of four is typical), rats swim as fast as they can to get from the start to the goal. This provides a measure of basic swimming ability and motivation to escape from water, and can be used to determine whether or not animals are motorically and motivationally equivalent across groups prior to MWM trials. An analysis of the average of these trials or use of the fastest trial provides assurance that MWM trials can be interpreted correctly.
Summary of the MWM
Spatial mapping versus working memory hypotheses and the data supporting each using the MWM has been reviewed in detail elsewhere5,42. There are many tests that have been used to assess these functions, of which the MWM is but one. However, the MWM has become an important, even dominant, method. As with all methods, the MWM has strengths and weaknesses; however, most of its perceived weaknesses arise from the use of mazes that are too small, protocols that do not adequately assess learning, failure to provide an appropriate interval between training and probe trials to assess reference memory, or lack control procedures to assess non-spatial factors. Despite this, the MWM has become more widely used than its predecessors (radial-arm maze, passive avoidance, T-mazes and their variations) since its introduction 25 years ago. This increased utilization arises because the effects on MWM performance after treatment have been more widely replicated than the effects observed with any other learning task, and the MWM is relatively straightforward to set up. There can be little doubt that the MWM has significantly advanced our understanding of the relationship between NMDA receptors, synaptic plasticity and learning43, and it continues to be used in new applications for the assessment of other types of learning. As use of the task has grown, so too have the number of methodological variations, some of which have extended its utility. The protocols described here provide guidance that can help users avoid the most common pitfalls.
The MWM is primarily a test of spatial learning and reference memory and that remains its principal strength. Detailed analyses have shown that rats can solve the task using a minimal set of cues that involve angular separation and distance from the tank wall29. Such data show that when properly configured and utilized, with the inclusion of appropriate control procedures, the MWM is a powerful technique for assessing spatial mapping. Appropriate modification of the basic protocol makes it a flexible tool that can be applied to probe spatial learning in more depth or to assess other forms of learning and memory.
Materials
Animals
-
Rats: Albino strains (such as Sprague–Dawley, Wistar, Fischer 344 and Lewis) learn well, despite reduced eye pigmentation, as do pigmented strains (such as Long–Evans and Listar).
-
Mice: Many labs report that C57BL mice are among the best performers in the MWM test44,45; however, many other strains and hybrids have been used (see GENETIC BACKGROUND).
-
Either rats or mice may be used, as described above.
Caution
Experimenters must comply with national regulations concerning animals and their use.
Equipment Setup
-
The Maze Morris' original maze was 130 cm in diameter1 but he later introduced a larger 214 cm diameter maze2. Thereafter, he refers to a maze of 200 cm diameter5 (see also ref. 46). Here, we will describe two apparatuses: one for rats and one for mice. Both are stainless-steel circular tanks. The one for rats is 210 cm in diameter and the one for mice is 122 cm in diameter. Both have sides that are 51 cm in height with non-reflective interior surfaces. For albino rats, the inside of the larger maze is painted black, whereas for C57BL mice, the inside of the smaller maze is painted white. Because the area of the surface increases with the square of the radius, larger tanks present a much more difficult spatial problem because of the greatly expanded search area to target ratio. The maze is typically mounted above the floor and plumbed with a drain line and valve to facilitate cleaning.
Caution
Commercially available swimming pools and troughs are often used as MWM tanks, but caution should be exercised with these as they often have prominent welded seams, corrugated surfaces or other features that provide proximal cues — that is, markings inside the tank. Such cues undermine the goal of the task, as the purpose is to test the animal's ability to navigate using distal cues. Having a tank that is professionally constructed with attention to unobtrusive seams and a uniform interior is worth the investment. Smooth interior walls also minimize the attempts of animals to climb the walls during early trials.
-
The platform The goal (platform or target) can be either square or circular and is usually 10 or 11 cm2 or 10–12 cm in diameter, and is typically submerged 1–2 cm below the surface, although with mice this depth is often reduced to 0.5–1 cm. We find it instructive to use smaller platform sizes for some phases of learning to increase task difficulty. Task difficulty is a function of the ratio of search area to target size. For example, a 210 cm diameter tank and a 10 cm2 platform, as we use, has a search area:target ratio of 346:1, whereas a 122 cm diameter tank and a 10 cm2 platform has a ratio of 117:1. These ratios work well for rats and mice, respectively. A reduction of the platform size to 5 cm2 increases this ratio to 1385:1 in the large tank and 468:1 in the small tank. We have previously compared the latencies for animals tested using a ratio of 346:1 (10 cm2 platform) or 1385:1 (5 cm2 platform) from the beginning of acquisition. We found that, although animals that are trained with the 10 cm2 platform learned the task to asymptotic performance within ∼20 trials, it took much longer using the 5 cm2 platform and performance never reached the same level as it did in the 10 cm2 platform trained group47. Therefore, starting out with a small platform makes the task too difficult. Smaller platforms are best reserved for later phases of testing, after the animal has learned the basic task with a larger platform. Research on the effect of tank diameter to target size is limited, but this ratio clearly is an important variable48.
Platforms are typically made of acrylic or PVC. There are several types, the most common being the use of an acrylic dowel or PVC pipe as the vertical post, with the platform mounted on the top (often with small holes or some other textured surface to provide traction). The base is usually a larger piece of acrylic that rests on the bottom of the pool and is large enough to provide stability so that it will not tip when the animal climbs on it and it resists movement when an animal bumps it. Another approach is to use a submerged acrylic disk with holes in it that the platform dowel fits in. This prevents erroneous positioning of the platform and makes it easier to precisely relocate the platform to other positions during different phases of testing. There are also platforms that can be raised and lowered (e.g., Atlantis On-Demand platforms, or by other means31). Most acrylic plastics have specific gravities <1.0, so these materials often require that weight be added. Selection of high-density acrylic avoids this problem.
-
Water A common concern is water temperature. It was originally suggested that rats need water to be warmer than typical ambient air temperatures that are found in most laboratories (19–22 °C), but this has not been widely borne out. Rats tested in water that has been equilibrated to ambient temperatures of 19–22 °C perform well and do not show evidence of significant fatigue or hypothermia at typical test ages49. However, the situation in mice may be different (see below).
-
Room configuration The maze should be placed in a room with ample surrounding visual cues. These distal cues can be intentional or can be endogenous to the room; however, it is critical that the cues are not moved during testing as these are the animal's navigational reference points for locating the goal, independent of start location. The distance of cues from the outside wall of the maze can also be important (see ref. 29). Because some labs are relatively barren, when in doubt it is prudent to add cues; there is no evidence that too many cues are a problem, but too few may be. It is useful to mount curtains around the maze. These may be used to position intentional cues should testing of the effects of cue rotation be desired, but the principal reason to have curtains is to obscure distal cues during cued learning trials (see above).
-
Experimenter Experimenter effects in the MWM principally involve animal handling and effects of experimenter visibility. Handling issues are the same as for any experiment that assesses behavior and will not be discussed here. The position of the experimenter can be handled in several ways: the experimenter can: (a) leave the room during testing; (b) stand behind a visual barrier; or (c) remain stationary in a constant location. The latter is the most common but it should not be forgotten that the experimenter then becomes one of the distal cues. Animals that are tested in different quadrants, especially in the distal versus the proximal half of the maze, sometimes show different latencies, depending on whether they are swimming towards or away from the experimenter50. This effect does not invalidate the test, but there should be awareness of it and outcomes should not be confounded by allowing an experimenter to move around as they wait for the animal to perform the task.
-
Lighting Room lighting should be indirect, especially if a video camera, computer and tracking software are used to record performance. Tracking software is sensitive to light reflections from the water surface and these are minimized by indirect lighting.
-
Tracking system There are many commercial vendors that provide tracking systems that can be used for the MWM. A non-exhaustive list includes HVS Image, San Diego Instruments, Accuscan, View Point, Clever Systems, Noldus, Columbus Instruments, Watermaze Software, Coulbourn Instruments, and others. As with any software system, it should be tested on-site before selection.
Procedure
Spatial acquisition
-
1
Place the animal in the desired start position in the maze, facing the tank wall. The animal is released into the water at water-level (not dropped). A timer or computer tracking program is started the moment that the animal is released.
-
2
Stop the timer when the animal reaches (touches) the platform (most animals immediately climb on the platform but there are exceptions36). A trial limit of 1 or 2 min per trial is standard; commonly, 2 min for rats and 1 min for mice. Animals not finding the platform within this time limit are either placed on the platform or guided to it.
-
3
Leave the animal on the platform during the inter-trial interval (ITI). Inter-trial intervals of 30 or 60 s were once used, but 15 s has become commonplace and produces good learning. However, longer ITIs are often used for mice, especially during the first test session and this improves learning25. This may be because mice are more prone to hypothermia-induced performance effects51. The objective in leaving the animal on the platform is to allow it to orient to its position in space and remember the position of the goal in relation to surrounding cues. However, this may not be necessary as animals may learn what they need during their navigation to the platform52. The latter investigators showed that rats learn the task just as well if the lights are on or off during the ITI when they remain on the platform.
-
4
Place the animal in the maze at a new start location and repeat the trial (Steps 1–3) until the animal has had the desired number of trials for that day. Animals are normally given multiple trials per day. The most common number is 4. As there are four principal start locations, this keeps the start positions balanced each day. With four trials per day, it takes 5–10 min per animal on the first day and progressively less time per animal each day thereafter. Other trial numbers that have been used are 1 (ref. 23), 2 (ref. 53), 3 (ref. 54), 5 (ref. 55), 6 (refs. 27,56,57), 8 (refs. 52,58), 10 (ref. 59) and 12 (refs. 60,61) trials per day. No large advantage of any one of these trial numbers has been convincingly demonstrated in rats. An alternative approach is to allow the animal 15 s on the platform, then remove it to its cage and test the second animal on Trial 1 and repeat this rotation until all animals have completed Trial 1, then repeat the process for subsequent trials. The latter method improves learning performance in mice51, but no comparable data exist for rats. There is little evidence to suggest large sensitivity differences as a function of daily trial number, but what evidence there is indicates that having fewer trials per day improves learning50. However, this issue has not been sufficiently explored to make definitive recommendations, and tests of trial number have often involved smaller tanks that may not generalize to larger mazes.
-
5
On subsequent days, repeat the trials. The number of days over which to repeat testing depends on the learning curve. With four trials per day, 5–6 days (20–24 trials) is typically sufficient in a 210 cm maze for rats or in a 122 cm maze for mice to reach asymptotic performance; however, as noted previously, if the task is made more difficult, then more days of testing may be required for animals to reach asymptotic performance. By contrast, rats in a 122 cm maze may approach asymptotic performance by day 2 or 3 (ref. 41).
Reference memory: probe trial
-
6
Remove the platform.
-
7
Place the animal in a novel start position in the maze, facing the tank wall — for example, 180° from the original platform position. We use a novel start position during the probe trial to ensure that its spatial preference is a reflection of the memory of the goal location rather than for a specific swim path (see Table 1).
-
8
Remove the animal after a fixed interval (usually 30 or 60 s). There is evidence that quadrant preference decreases after the first 30 s (ref. 41), hence 30 s is recommended. The object of the probe trial is to determine whether or not the animal remembers where the platform was located. Indications of such memory include number of platform-site crossovers, time and distance spent in the target quadrant compared with the other quadrants, time in a pre-defined annulus surrounding the target that is larger than the target itself, average distance to the target site, angle (bearing) to the target site, latency to first target-site crossover, and mean search difference and mean zone difference scores (see ref. 62 for a description of the latter measures). Percent time or percent distance in the target quadrant is used most frequently.
Critical Step
Probe trials that are intended to assess reference memory should not be administered shortly after the last learning trial, as this measures recent rather than long-term recall. If an immediate probe trial is given, it should not be a substitute for one given at least 24 h later.
Spatial reversal
-
9
If spatial reversal tests are desired, relocate the platform to another quadrant (usually the opposite one) and administer another set of four trials per day for 5 days (as described in Steps 1–4).
-
10
At the end of the reversal phase, perform a reversal probe trial, as described in Steps 6–8.
Troubleshooting
The most common problem is using an improper tank to platform size combination. Many tanks are too small or are at the lower boundary of what constitutes an optimal task for assessing spatial learning. Smaller tanks allow the animal to use other strategies (such as swimming a fixed distance from the wall) to solve the task, while making minimal use of distal cues. Unfortunately, if the tank is too small, animals will learn and the experimenter will not be aware that the animal is using non-distal cues to find the platform and nothing will appear amiss. For this reason, selection of the proper tank size from the outset is the best method for obtaining valid spatial learning.
Using untreated, adult, male, Long–Evans rats, the effect of tank size on performance can be seen in Figure 3. The small tank is 122 cm in diameter and the large tank is 210 cm in diameter. Water temperature was the same for both groups, as was platform size (10 cm2). Both groups received the same number of trials (four per day), in the same order and to the same goal position, on the same days in the same room. The tanks were identically constructed and painted, and the only difference was diameter. For the small tank trials, the smaller perimeter was placed inside the larger one. As can be seen, the small tank group learned the task well; in fact, they learned it so rapidly that there is almost no learning curve. Basically, the small tank group approached asymptotic performance on the second day. By contrast, the large tank group showed a steady improvement in learning up to day 4 and reached asymptotic performance on day 5. It is apparent which curve is better for assessing learning; hence, a 122 cm tank is not recommended for rats.

Rats were given four trials per day and data represent the mean ± s.e.m. of blocks of four trials. Data are latency (s) to reach the goal, using identical sequences of start locations. The small tank was 122 cm in diameter and the large tank was 210 cm in diameter. All testing conditions and trials were identical for the two groups. The small tank perimeter was placed inside the larger tank so that all cues were identical. n = 10 per group (males).
On the other hand, a 122 cm tank is the most common size that is used for mice. Transgenic, knock-out and knock-in mice are routinely assessed in the MWM for spatial learning. There are ample data that, in mice, good learning curves can be obtained using a 122 cm diameter tank. This may be because this size tank is scaled more to the body size of a mouse or because mice find spatial tasks more difficult than rats and, therefore, a smaller search area is beneficial.
Different mouse strains vary widely in performance during the MWM test. One strain that performs reasonably well is C57BL (ref. 57). Regardless of vendor, C57BL mice are fairly good swimmers and respond to being placed in water with an appropriate swim-search response. Some strains float or turn in circles and cannot be used.
Genetic background
In rats, Sprague–Dawley, Wistar, Long–Evans, Fischer 344, ACI, Dark Agouti and Lewis have all been shown to learn when tested in the MWM.
In mice, the results are consistent that C57BL learn in the MWM, but labs have differed in what they report for other strains. For example, 129S strains in which gene targeting is often performed, have an increased incidence of floating and erratic performance but this may be a function of substrain, as 129S2/SvlmJ mice are reported to perform satisfactorily in the MWM45. BALB mice are reported to perform well by some45 in the MWM and poorly by others44.
A factor analysis that was designed to locate clustered determinants of MWM learning across multiple mouse strains and a large number of mice (∼1,500) has shown that three factors account for most of the variance in MWM performance63,64. The largest factor in mice (48%) was termed thigmotaxis and represents the tendency to swim or float near the tank wall; the second largest factor was termed passivity (20%) and represents slow or non-swimming; and the third factor was memory or goal-directed behavior (13%). The outcome of such an analysis is a function of the factors measured, so different measurements would generate a different factor analysis. Nevertheless, this analysis, which included many important variables of MWM performance, shows that much of the variance in performance is due to factors that are unrelated to learning and memory in mice, hence use of appropriate controls is especially important in mouse studies to arrive at a correct interpretation of results.
Non-performers
Non-performers are rarely encountered in rats, but are seen in mice. Among C57BL mice, non-performers in the MWM are less common than in most other strains. However, some genetic modifications can cause mutant animals to not perform the task. These cases are relatively obvious. However, having a subset of mice that do not search for the goal, because of mixed genetic background or the genetic manipulation under investigation, occurs with some frequency. In such situations, the problem becomes how to handle such cases. The most common problem is that some of the mice float and appear unmotivated to escape. Different methods have been tried to induce non-performers to swim, but these approaches often involve experimenter interventions, such as startling the mouse with a loud noise, prodding it with an object or pre-wetting the fur to reduce buoyancy. Mice are buoyant because their fur coat contains water-repellant oils that allow them to remain floating for minutes at a time. Testing mice in 20–22 °C water generally reduces the frequency of floating. Nonetheless, caution should be used in allowing experimenter interventions. The problem is that interventions are difficult to apply uniformly and may inadvertently introduce bias. If the experimental group has a higher or lower floating frequency or the treatment interacts with startle or other factors, the results could be confounded. A better approach is use of a standard rule that does not involve intervention. Our approach is that if a mouse floats, it is allowed to remain in the water up to the limit of the trial. If the animal fails to swim, it is removed and the next animal is tested. When the other animals in the session are finished, the floater is given a second trial. If it swims, it receives its full trial sequence, with the first trial being excluded from the data. If it floats for the second trial, it is returned to its cage and tested again the next day. If it now swims, it proceeds though the test, staggered one day behind its cohort. If it fails to swim, it is taken out of the test order, as on day 1, until the other animals in the session are tested and is then given one final trial. If it fails to swim during this fourth attempt, it is removed from the experiment. At the conclusion of the experiment, the frequency of floaters in each group is statistically compared to determine whether or not the experimental manipulation significantly altered the frequency of non-performers compared with the wild type. If it does, then the MWM data on those that did swim must be interpreted with caution in light of the possibility of selection bias.
Timing
Each trial is limited to 2 min in rats and 1 min in mice, with an ITI of 15 s and four trials per day. Therefore, it takes no longer than 9 min to complete a learning session. As animals learn, the time per day steadily decreases. Probe trials on the last day are only 30 s. For any phase of testing, 6 days is common: 5 learning days and 1 probe day. The first phase is often cued, followed by acquisition, then reversal, double-reversal, working memory (matching to sample), or other procedures.
Anticipated results
Learning trials
Dependent measures of performance on learning trials in the MWM begin with latency (time from start to goal). Latency can be obtained with or without tracking software. Path length is also widely used and has the advantage of being immune to swimming speed. Although path-length determinations have been carried out without the use of a tracking program, these methods are labor-intensive and such data are more easily captured with a tracking program. Another useful index is cumulative distance from the platform; most tracking programs generate this measure as the distance from the platform measured every second or every few milliseconds. Cumulative distance has been suggested to be one of the better measures of spatial learning ability54. First bearing and/or mean directionality, which are similar but not identical, measure the animal's swim angle at the start of the trial relative to a direct line from the start to the goal. Angle measurements appear to be sensitive to many types of experimental treatments. In our experience, latency, path length and cumulative distance are highly correlated, whereas first bearing has a lower correlation with these first three measures (Table 4). This is logical as the first three measures all reflect aspects of performance across the length of the trial, whereas first bearing is a snapshot of the animal's performance at one particular point during the trial (at the beginning). Other measures are time or distance in the target annulus (the space between two concentric circles, defined by lines touching the outside and inside edges of the platform, respectively — not to be confused with Morris' use of the term 'annulus crossing', by which he means crossings of a zone surrounding where the platform had been) compared with inner or outer annuli (the latter being a measure of thigmotaxis), number of direct swims (number of paths within a preset corridor from the start to the goal) or circle swims (a swim trajectory that approximates a decreasing radius arc of a circle from the start point to the hidden platform and circling is less than 360°), jump-offs, deflections, swimovers, and others. These other measures can be important especially when testing animals that have recently been administered a drug that can disrupt sensorimotor function36,37.
Probe trials
Site crossings as an index of memory has the limitation that it is variable and often has a low frequency of occurrence, especially if the target is reduced in size or the tank diameter is large. Furthermore, depending on the tracking software, crossover undercounting may occur. We find that the average distance to the target site, time or distance in the target quadrant and first bearing provide more robust measures of memory than exact site crossings. Others report success with target annulus crossovers, where the annulus is a circular zone surrounding and larger than the platform itself.
Data analysis
Acquisition trials are generally averaged in blocks of four and plotted as block means (± s.e.m.), or if larger numbers of trials are used then there may be two or more blocks of trials plotted per day. However, the experimenter should also examine the data by trial to ensure that learning is occurring within each daily test session. Unless an unusual pattern is seen, plotting the data in daily four-trial blocks usually accurately represents the learning process. When daily trials are plotted, long latencies are generally seen during Trial 1 and Trial 2 of day 1, with improvement during Trial 3 and Trial 4. The next day, performance will begin with Trial 1 being longer than Trial 4 of the preceding day, and then performance improving to exceed the performance on Trial 4 of day 1. This saw-tooth pattern repeats on each successive day of testing, with an overall shortening or downward stair-step pattern of performance across days. Plotting the data by blocks of trials smoothes the learning curve and this line is the most widely used index of spatial learning, although other indices have been used (for example, ref. 54).
Data summary and analysis
Group means for each dependent variable should be calculated per trial and per block of trials. Trials 1 and 2 of day 1 of testing should be examined to ensure that the groups do not start the test at different performance levels. The data on the first one or two trials should be compared using a test with high detection power, such as a t-test or analysis of variance (ANOVA) if there are more than two groups. If the groups differ significantly during early trials, it is a warning that the groups entered the test with preexisting differences that may prevent or compromise the interpretation of the learning curves or retention trials in terms of spatial learning and reference memory. If the groups are comparable, then the data are typically plotted and analyzed in blocks of trials in order to stabilize the means. Data are generally analyzed by ANOVA. The factors are usually genotype or treatment group and test day. Some experiments will include additional factors, such as sex. Group is generally a 'between'-subject factor and day a 'within'-subject factor in the ANOVA model, although in developmental studies using split-litter designs the factors of group, day and sex may all be within/matching factors. Using statistical programs such as SAS, MWM data may be analyzed using Proc GLM (general linear model) or Proc Mixed. GLM split-plot models can accommodate one or more 'between' and one or more 'within' factors; however, one must be cautious when using GLM models for repeated measure ('within') variables. If there are only two levels of the 'within' factors, the GLM ANOVA solution is precise, but in the MWM there are usually more than two days of testing. With more than two levels of the repeated measure factor, the significance of the 'within' and 'between' x 'within' factor F-ratios can be distorted. This is because GLM requires that the data meet the assumption of compound symmetry — that is, that the correlations within the variance-covariance matrix of the repeated factor, say day, be exactly the same at all distances between measurements. For example, the correlation between day 1 and day 2 must be the same as the correlation between day 1 and day 3 or between day 2 and day 3. This assumption is almost never met by learning data because variances change as a function of day as learning proceeds. As animals learn the task, their performance become less variable, hence the variances decrease across days and the correlations change. There is no exact test for compound symmetry, but there is a more stringent test for sphericity. If GLM is used, it is advisable that the covariance matrix test for sphericity be used. If the data are significantly non-spherical, there are adjusted F-ratios that can be used to at least partially correct the F-tests so that significance levels are less likely to be overestimated. The two most common methods of adjusting F-ratios are those of Greenhouse–Geisser and Huyhn–Feldt.
Another approach is to use Proc Mixed ANOVA models (SAS). Proc Mixed ANOVA models provide a more precise way of handling the problem of non-symmetry of the covariance matrix, but Proc Mixed is limited in that it cannot handle more than two 'within' dimensions in the model. However, Proc Mixed has some significant advantages that make it worthwhile. Proc Mixed provides a set of different covariance models that can be fitted to the data. These different models can be tested against the data and compared using best-fit statistics provided by Proc Mixed. In our experience, the AR (1) (autoregressive (1)) covariance model often provides the best fit for MWM learning variables, such as latency, path length and cumulative distance. Regardless of which covariance model best fits the data, the best fit model can be chosen and the data can be analyzed to obtain more precise F-tests than with Proc GLM. Probe data often have no repeated measure component and, therefore, do not require Proc Mixed. Proc Mixed also provides a 'slice' ANOVA option, such that if an interaction is obtained between the factors of group and day, the slice ANOVAs will provide tests of the group effect on each day in order to localize which days show group differences. From this, individual group comparisons (post hoc tests) can be made between the treatment groups on selected days; significant interactions or group main effects require the typical considerations of controlling for multiple comparisons. In SAS, Proc Multtest provides group comparison tests that provide different levels of protection against type I errors. The step-down Bonferonni is one such test that we commonly use.
References
Morris, R.G.M. Spatial localization does not require the presence of local cues. Learn. Motiv. 12, 239–260 (1981).
Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 11, 47–60 (1984).
Stewart, C.A. & Morris, R.G.M. The watermaze. in Behavioural Neuroscience, Volume I, A Practical Approach (ed. Sahgal, A.) 107–122 (IRL Press at Oxford University Press, Oxford, 1993).
Kallai, J., Makany, T., Karadi, K. & Jacobs, W.J. Spatial orientation strategies in Morris-type virtual water task for humans. Behav. Brain Res. 159, 187–196 (2005).
Morris, R.G.M. An attempt to dissociate 'spatial-mapping' and 'working-memory' theories of hippocampal function. in Neurobiology of the Hippocampus (ed. Seifert, W.) 405–432 (Academic Press, New York, 1993).
Cravens, R.W. Effects of maternal undernutrition on offspring behavior: Incentive value of a food reward and ability to escape from water. Dev. Psychobiol. 7, 61–69 (1974).
Morris, R.G., Garrud, P., Rawlins, J.N. & O'Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature 297, 681–683 (1982).
Fitzgerald, L.W. & Dokla, C.P. Morris water task impairment and hypoactivity following cysteamine-induced reductions of somatostatin-like immunoreactivity. Brain Res. 505, 246–250 (1989).
Brandeis, R., Brandys, Y. & Yehuda, S. The use of the Morris water maze in the study of memory and learning. Int. J. Neurosci. 48, 29–69 (1989).
D'Hooge, R. & De Deyn, P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res. Rev. 36, 60–90 (2001).
McNamara, R.K. & Skelton, R.W. The neuropharmacological and neurochemical basis of place learning in the Morris water maze. Brain Res. Rev. 18, 33–49 (1993).
Jeffery, K.J. & Morris, R.G.M. Cumulative long-term potentiation in the rat dentate gyrus correlates with, but does not modify, performance in the water maze. Hippocampus 3, 133–140 (1993).
Moser, E.I., Krobert, K.A., Moser, M.-B. & Morris, R.G.M. Impaired spatial learning after saturation of long-term potentiation. Science 281, 2038–2042 (1998).
Morris, R.G.M., Anderson, E., Lynch, G.S. & Baudry, M. Selective impairment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate antagonist, AP5. Nature 329, 774–776 (1986).
Bannerman, D.M., Good, M.A., Butcher, S.P., Ramsay, M. & Morris, R.G.M. Distinct components of spatial learning revealed by prior training and NMDA receptor blockade. Nature 378, 182–186 (1995).
Vorhees, C.V. et al. Adult learning deficits after neonatal exposure to D-methamphetamine: Selective effects on spatial navigation and memory. J. Neurosci. 20, 4732–4739 (2000).
Broening, H.W., Morford, L.L., Inman-Wood, S.L., Fukumura, M. & Vorhees, C.V. 3,4-methylenedioxymethamphetamine (ecstasy) induced learning and memory impairments depend on the age of exposure during early development. J. Neurosci. 21, 3228–3235 (2001).
Vorhees, C.V., Reed, T.M., Skelton, M.R. & Williams, M.T. Exposure to 3,4-methylenedioxymethamphetamine (MDMA) on postnatal days 11-20 induces reference but not working memory deficits in the Morris water maze in rats: implications of prior learning. Int. J. Dev. Neurosci. 22, 247–259 (2004).
Williams, M.T. et al. Developmental d-methamphetamine treatment selectively induces spatial navigation impairments in reference memory in the Morris water maze while sparing working memory. Synapse 48, 138–148 (2003).
Morford, L.L., Inman-Wood, S.L., Gudelsky, G.A., Williams, M.T. & Vorhees, C.V. Impaired spatial and sequential learning in rats treated neonatally with d-fenfluramine. Eur. J. Neurosci. 16, 491–500 (2002).
Ehrman, L.A. et al. Phosphodiesterase 1B modulates the effects of methamphetamine on locomotor activity and spatial learning through a DARPP-32-dependent pathway: Evidence from PDE1B-DARPP32 double-knockout mice. Genes Brain Behav. (in the press).
Silva, A.J., Paylor, R., Wehner, J.M. & Tonegawa, S. Impaired spatial learning in α-calcium-calmodulin kinase II mutant mice. Science 257, 206–211 (1992).
Bourtchuladze, R. et al. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 79, 59–68 (1994).
Cho, Y.H., Giese, K.P., Tanila, H., Silva, A.J. & Eichenbaum, H. Abnormal hippocampal spatial representations in αCaMKIIT286A and CREBαΔ- mice. Science 279, 867–869 (1998).
Upchurch, M. & Wehner, J.M. Effects of N-methyl-D-aspartate antagonism on spatial learning in mice. Psychopharmacology 100, 214 (1990).
Grant, S.G.N. et al. Impaired long-term potentiation, spatial learning, and hippocampal development in fyn mutant mice. Science 258, 1903–1910 (1992).
Limback-Stokin, K., Korzus, E., Nagaoka-Yasuda, R. & Mayford, M. Nuclear calcium/calmodulin regulates memory consolidation. J. Neurosci. 24, 10858–10867 (2004).
Tang, Y.-P. et al. Genetic enhancement of learning and memory in mice. Nature 401, 63–69 (1999).
Maurer, R. & Derivaz, V. Rats in a transparent morris water maze use elemental and configural geometry of landmarks as well as distance to the pool wall. Spatial Cogn. Comput. 2, 135–156 (2000).
Kim, J.J., Lee, H.J., Han, J.-S. & Packard, M.G. Amygdala is critical for stress-induced modulation of hippocampal long-term potentiation and learning. J. Neurosci. 21, 5222–5228 (2001).
Markowska, A.L., Long, J.M., Johnson, C.T. & Olton, D.S. Variable-interval probe test as a tool for repeated measurements of spatial memory in the water maze. Behav. Neurosci. 107, 627–632 (1993).
Sutherland, R.J., Chew, G.L., Baker, J.C. & Linggard, R.C. Some limitations on the use of distal cues in place navigation by rats. Psychobiology 15, 48–57 (1987).
Williams, M.T. et al. Developmental 3,4-methylenedioxymethamphetamine (MDMA) impairs sequential and spatial but not cued learning independent of growth, litter effects, or injection stress. Brain Res. 968, 89–101 (2003).
Saucier, D. & Cain, D.P. Spatial learning without NMDA receptor-dependent long-term potentiation. Nature 378, 186–189 (1995).
Saucier, D., Hargreaves, E.L., Boon, F., Vanderwolf, C.H. & Cain, D.P. Detailed behavioral analysis of water maze acquisition under systemic NMDA or muscarinic antagonism: Nonspatial pretraining eliminates spatial learning deficits. Behav. Neurosci. 110, 103–116 (1996).
Cain, D.P., Saucier, D., Hall, J., Hargreaves, E.L. & Boon, F. Detailed behavioral analysis of water maze acquisition under APV or CNQX: Contribution of sensorimotor disturbances to drug-induced acquisition deficits. Behav. Neurosci. 110, 86–102 (1996).
Cain, D.P. & Saucier, D. The neuroscience of spatial navigation: Focus on behavior yields advances. Rev. Neurosci. 7, 215–231 (1996).
Hoh, T.E. & Cain, D.P. Fractionating the nonspatial pretraining effect in the water maze task. Behav. Neurosci. 111, 1285–1291 (1997).
Williams, M.T., Vorhees, C.V., Boon, F., Saber, A.J. & Cain, D.P. Methamphetamine exposure from postnatal days 11 to 20 causes impairments in both behavioral strategies and spatial learning in adult rats. Brain Res. 958, 312–321 (2002).
Whishaw, I.Q. Posterior neocortical (visual cortex) lesions in the rat impair matching-to-place navigation in a swimming pool: a reevaluation of cortical contributions to spatial behavior using a new assessment of spatial versus non-spatial behavior. Behav. Brain Res. 155, 177–184 (2004).
Blokland, A., Geraerts, E. & Been, M. A detailed analysis of rats' spatial memory in a probe trial of a Morris task. Behav. Brain Res. 154, 71–75 (2004).
Morris, R.G.M., Hagan, J.J. & Rawlins, J.N.P. Allocentric spatial learning by hippocampectomized rats: A further test of the 'spatial mapping' and 'working memory' theories of hippocampal function. Quart. J. Exp. Psychol. 38B, 365–395 (1986).
Morris, R.G.M., Davis, S. & Butcher, S.P. Hippocampal synaptic plasticity and NMDA receptors: A role in information storage? Phil. Trans. R. Soc. Lond. 329, 187–204 (1990).
Upchurch, M. & Wehner, J.M. Differences between inbred strains of mice in Morris water maze performance. Behav. Genet. 18, 55–68 (1988).
Wahlsten, D., Cooper, S.F. & Crabbe, J.C. Different rankings of inbred mouse strains on the Morris maze and a refined 4-arm water escape task. Behav. Brain Res. 165, 36–51 (2005).
Wenk, G.L. Assessment of spatial memory using the radial arm and Morris water maze. (eds. Crawley, J.N. et al.) Unit 8.5A (Wiley Interscience, New York, 2004).
Williams, M.T., Moran, M.S. & Vorhees, C.V. Behavioral and growth effects induced by low dose methamphetamine administration during the neonatal period in rats. Int. J. Dev. Neurosci. 22, 273–283 (2004).
Mactutus, C.F. & Booze, R.M. Accuracy of spatial navigation: The role of platform and tank size. Soc. Neurosci. Abst. 20, 1014 (1994).
Lindner, M.D. & Ribkoff, V.K. Relationship between performance in the Morris water task, visual acuity, and thermoregulatory function in aged F-344 rats. Behav. Brain Res. 45, 45–55 (1991).
Hodges, H. Maze procedures: The radial-arm and water maze compared. Cog. Brain Res. 3, 167–181 (1996).
Iivonen, H., Nurminen, L., Harri, M., Tanila, H. & Puolivali, J. Hypothermia in mice tested in Morris water maze. Behav. Brain Res. 141, 207–213 (2003).
Devan, B.D., Blank, G.S. & Petri, H.L. Place navigation in the Morris water task: Effects of reducted platform interval lighting and pseudorandom plaform positioning. Psychobiology 20, 120–126 (1992).
Otnaess, M.K., Brun, V.H., Moser, M.-B. & Moser, E.I. Pretraining prevents spatial learning impairment after saturation of hippocampal long-term potentiation. J. Neurosci. 19 (RC49): 1–5 (1999).
Gallagher, M., Burwell, R. & Burchinal, M. Severity of spatial learning impairment in aging: Development of a learning index for performance in the Morris water maze. Behav. Neurosci. 107, 618–626 (1993).
Burwell, R.D., Saddoris, M.P., Bucci, D.J. & Wiig, K.A. Corticohippocampal contributions to spatial and contextual learning. J. Neurosci. 24, 3826–3836 (2004).
Holscher, C. Stress impairs performance in spatial watermaze learning tasks. Behav. Brain Res. 100, 225–235 (1999).
Clapcote, S.J., Lazar, N.L., Bechard, A.R., Wood, G.A. & Roder, J.C. NIH Swiss and Black Swiss mice have retinal degeneration and performance deficits in cognitive tests. Comp. Med. 55, 310–316 (2005).
Devan, B.D., McDonald, R.J. & White, N.M. Effects of medial and lateral caudate-putamen lesions on place- and cue-guided behaviors in the water maze: relation to thigmotaxis. Behav. Brain Res. 100, 5–14 (1999).
Rauch, T.M., Welch, D.I. & Gallego, L. Hyperthermia impairs retrieval of an overtrained spatial task in the Morris water maze. Behav. Neural Biol. 52, 321–330 (1989).
Giese, K.P., Fedorov, N.B., Filipkowski, R.K. & Silva, A.J. Autophosphorylation at Thr286 of the α calcium-calmodulin kinase II in LTP and learning. Science 279, 870–873 (1998).
Guzowski, J.F., Setlow, B., Wagner, E.K. & McGaugh, J.L. Experience-dependent gene expression in the rat hippocampus after spatial learning: A comparison of the immediate-early genes Arc, c-fos, and zif268. J. Neurosci. 21, 5089–5098 (2001).
Brown, R.W. et al. Adulthood nicotine treatment alleviates behavioural impairments in rats neonatally treated with quinpirole: possible roles of acetylcholine function and neurotrophic factor expression. Eur. J. Neurosci. 19, 1634–1642 (2004).
Wolfer, D.P., Stagljar-Bozicevic, M., Errington, M.L. & Lipp, H.-P. Spatial memory and learning in transgenic mice: Fact or artifact? News Physiol. Sci. 13, 118–123 (1998).
Lipp, H.P. & Wolfer, D.P. Genetically modified mice and cognition. Curr. Opin. Neurobiol. 8, 272–280 (1998).
Acknowledgements
The authors acknowledge the support of National Institutes of Health grants DA006733, DA021394, DA014269 and ES007051.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
About this article
Cite this article
Vorhees, C., Williams, M. Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nat Protoc 1, 848–858 (2006). https://doi.org/10.1038/nprot.2006.116
Published:
Issue Date:
DOI: https://doi.org/10.1038/nprot.2006.116
This article is cited by
-
Cognitive decline, Aβ pathology, and blood–brain barrier function in aged 5xFAD mice
Fluids and Barriers of the CNS (2024)
-
Puerarin attenuates valproate-induced features of ASD in male mice via regulating Slc7a11-dependent ferroptosis
Neuropsychopharmacology (2024)
-
Hexahydrocurcumin attenuated demyelination and improved cognitive impairment in chronic cerebral hypoperfusion rats
Inflammopharmacology (2024)
-
Urolithin A Prevents Sleep-deprivation-induced Neuroinflammation and Mitochondrial Dysfunction in Young and Aged Mice
Molecular Neurobiology (2024)
-
MSC-Derived Extracellular Vesicles Alleviate NLRP3/GSDMD-Mediated Neuroinflammation in Mouse Model of Sporadic Alzheimer’s Disease
Molecular Neurobiology (2024)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
