
Key Points
-
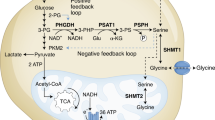
Serine is a non-essential amino acid that can be taken up by the cell or synthesized de novo from glycolytic intermediates through the serine synthesis pathway (SSP). Serine is used for many biosynthetic pathways, including the one-carbon cycle that supports nucleotide synthesis, methylation reactions and antioxidant defence.
-
Although one-carbon metabolism enzymes are expressed in both the mitochondria and cytosol, most cells generate formate in the mitochondria for use in cytosolic nucleotide synthesis. This directionality in the pathway can also provide mitochondrial NADH, NADPH and ATP.
-
Some cancer cells show amplification of the SSP enzymes, and their survival depends on the sustained expression of these enzymes, even under serine-fed conditions. This seems to reflect a requirement for serine synthesis to maintain nucleotide synthesis.
-
Cancer cells without amplification of the SSP tend to depend on exogenous serine for optimal growth. The switch to de novo serine synthesis under serine starvation leads to increased oxidative stress, which must be managed for successful adaptation.
-
Several oncogenes and tumour suppressor genes can regulate the serine synthesis and one-carbon cycle enzymes, with evidence that changes in their expression and activity can contribute to malignant development and progression.
-
Therapeutic approaches are being developed based on limiting exogenous serine or targeting serine synthesis or metabolism enzymes.
Abstract
The non-essential amino acid serine supports several metabolic processes that are crucial for the growth and survival of proliferating cells, including protein, amino acid and glutathione synthesis. As an important one-carbon donor to the folate cycle, serine contributes to nucleotide synthesis, methylation reactions and the generation of NADPH for antioxidant defence. Many cancer cells are highly dependent on serine, a trait that provides several novel therapeutic opportunities, either through the inhibition of de novo serine synthesis or by limiting the availability or uptake of exogenous serine.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

References
Phillips, M. M., Sheaff, M. T. & Szlosarek, P. W. Targeting arginine-dependent cancers with arginine-degrading enzymes: opportunities and challenges. Cancer Res. Treat. 45, 251–262 (2013).
Altman, B. J., Stine, Z. E. & Dang, C. V. From Krebs to clinic: glutamine metabolism to cancer therapy. Nat. Rev. Cancer https://dx.doi.org/10.1038/nrc.2016.71 (2016).
Maddocks, O. D. et al. Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature 493, 542–546 (2013). This paper describes how the tumour suppressor p53 helps cells to adapt to serine starvation, and a therapeutic benefit of dietary depletion of serine in a mouse xenograft model.
Kalhan, S. C. & Hanson, R. W. Resurgence of serine: an often neglected but indispensable amino acid. J. Biol. Chem. 287, 19786–19791 (2012).
Furuya, S. An essential role for de novo biosynthesis of l-serine in CNS development. Asia Pac. J. Clin. Nutr. 17 (Suppl. 1), 312–315 (2008).
De Koning, T. J. & Klomp, L. W. Serine-deficiency syndromes. Curr. Opin. Neurol. 17, 197–204 (2004).
Yoshida, K. et al. Targeted disruption of the mouse 3-phosphoglycerate dehydrogenase gene causes severe neurodevelopmental defects and results in embryonic lethality. J. Biol. Chem. 279, 3573–3577 (2004).
Jaeken, J. et al. 3-Phosphoglycerate dehydrogenase deficiency and 3-phosphoserine phosphatase deficiency: inborn errors of serine biosynthesis. J. Inherit. Metab. Dis. 19, 223–226 (1996).
Hart, C. E. et al. Phosphoserine aminotransferase deficiency: a novel disorder of the serine biosynthesis pathway. Am. J. Hum. Genet. 80, 931–937 (2007).
Jain, M. et al. Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation. Science 336, 1040–1044 (2012). A large-scale analysis of nutrient uptake in cancer cells shows avid consumption of serine and demonstrates a role for folate cycle enzymes in cancer development.
Kalhan, S. C. et al. Metabolic and genomic response to dietary isocaloric protein restriction in the rat. J. Biol. Chem. 286, 5266–5277 (2011).
Sun, L. et al. cMyc-mediated activation of serine biosynthesis pathway is critical for cancer progression under nutrient deprivation conditions. Cell Res. 25, 429–444 (2015).
Chaneton, B. et al. Serine is a natural ligand and allosteric activator of pyruvate kinase M2. Nature 491, 458–462 (2012).
Ye, J. et al. Pyruvate kinase M2 promotes de novo serine synthesis to sustain mTORC1 activity and cell proliferation. Proc. Natl Acad. Sci. USA 109, 6904–6909 (2012). References 13 and 14 describe the consequences of decreased intracellular serine in the inhibition of PKM2 and promotion of the SSP.
Hitosugi, T. et al. Phosphoglycerate mutase 1 coordinates glycolysis and biosynthesis to promote tumor growth. Cancer Cell 22, 585–600 (2012).
Kung, C. et al. Small molecule activation of PKM2 in cancer cells induces serine auxotrophy. Chem. Biol. 19, 1187–1198 (2012).
Davis, S. R. et al. Tracer-derived total and folate-dependent homocysteine remethylation and synthesis rates in humans indicate that serine is the main one-carbon donor. Am. J. Physiol. Endocrinol. Metab. 286, E272–E279 (2004).
Allen, R. W. & Moskowitz, M. Arrest of cell growth in the G1 phase of the cell cycle by serine deprivation. Exp. Cell Res. 116, 127–137 (1978).
Rowe, P. B., Sauer, D., Fahey, D., Craig, G. & McCairns, E. One-carbon metabolism in lectin-activated human lymphocytes. Arch. Biochem. Biophys. 236, 277–288 (1985).
Brosnan, M. E., MacMillan, L., Stevens, J. R. & Brosnan, J. T. Division of labour: how does folate metabolism partition between one-carbon metabolism and amino acid oxidation? Biochem. J. 472, 135–146 (2015).
Tibbetts, A. S. & Appling, D. R. Compartmentalization of mammalian folate-mediated one-carbon metabolism. Annu. Rev. Nutr. 30, 57–81 (2010). An authoritative review of the compartmentalization of the reactions in one-carbon metabolism.
Vazquez, A., Markert, E. K. & Oltvai, Z. N. Serine biosynthesis with one carbon catabolism and the glycine cleavage system represents a novel pathway for ATP generation. PLoS ONE 6, e25881 (2011).
Tedeschi, P. M. et al. Contribution of serine, folate and glycine metabolism to the ATP, NADPH and purine requirements of cancer cells. Cell Death Dis. 4, e877 (2013).
Ducker, G. S. et al. Reversal of cytosolic one-carbon flux compensates for loss of the mitochondrial folate pathway. Cell Metab. 23, 1140–1153 (2016). This study shows direct evidence for the plasticity of the folate cycle.
Nikkanen, J. et al. Mitochondrial DNA replication defects disturb cellular dNTP pools and remodel one-carbon metabolism. Cell Metab. 23, 635–648 (2016).
Pasternack, L. B., Laude, D. A. Jr & Appling, D. R. 13C NMR detection of folate-mediated serine and glycine synthesis in vivo in Saccharomyces cerevisiae. Biochemistry 31, 8713–8719 (1992).
Herbig, K. et al. Cytoplasmic serine hydroxymethyltransferase mediates competition between folate-dependent deoxyribonucleotide and S-adenosylmethionine biosyntheses. J. Biol. Chem. 277, 38381–38389 (2002).
Gregory, J. F. 3rd et al. Primed, constant infusion with [2H3]serine allows in vivo kinetic measurement of serine turnover, homocysteine remethylation, and transsulfuration processes in human one-carbon metabolism. Am. J. Clin. Nutr. 72, 1535–1541 (2000).
Fan, J. et al. Quantitative flux analysis reveals folate-dependent NADPH production. Nature 510, 298–302 (2014). This study provides evidence that in some cells, one-carbon metabolism can be an important source of NADPH.
Tucker, E. J. et al. Mutations in MTFMT underlie a human disorder of formylation causing impaired mitochondrial translation. Cell Metab. 14, 428–434 (2011).
Nilsson, R. et al. Metabolic enzyme expression highlights a key role for MTHFD2 and the mitochondrial folate pathway in cancer. Nat. Commun. 5, 3128 (2014).
Lewis, C. A. et al. Tracing compartmentalized NADPH metabolism in the cytosol and mitochondria of mammalian cells. Mol. Cell 55, 253–263 (2014). An elegant use of mutant forms of IDH to analyse the compartmentalization of NADPH production.
Zhao, H., French, J. B., Fang, Y. & Benkovic, S. J. The purinosome, a multi-protein complex involved in the de novo biosynthesis of purines in humans. Chem. Commun. (Camb.) 49, 4444–4452 (2013).
Chan, C. Y. et al. Purinosome formation as a function of the cell cycle. Proc. Natl Acad. Sci. USA 112, 1368–1373 (2015).
French, J. B. et al. Spatial colocalization and functional link of purinosomes with mitochondria. Science 351, 733–737 (2016).
Girgis, S. et al. Molecular cloning, characterization and alternative splicing of the human cytoplasmic serine hydroxymethyltransferase gene. Gene 210, 315–324 (1998).
Suh, J. R., Herbig, A. K. & Stover, P. J. New perspectives on folate catabolism. Annu. Rev. Nutr. 21, 255–282 (2001).
Field, M. S., Kamynina, E. & Stover, P. J. MTHFD1 regulates nuclear de novo thymidylate biosynthesis and genome stability. Biochimie 126, 27–30 (2016).
Bao, X. R. et al. Mitochondrial dysfunction remodels one-carbon metabolism in human cells. eLife 5, e10575 (2016).
Stincone, A. et al. The return of metabolism: biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. Camb. Philos. Soc. 90, 927–963 (2015).
Liu, L. et al. Malic enzyme tracers reveal hypoxia-induced switch in adipocyte NADPH pathway usage. Nat. Chem. Biol. 12, 345–352 (2016).
Bolusani, S. et al. Mammalian MTHFD2L encodes a mitochondrial methylenetetrahydrofolate dehydrogenase isozyme expressed in adult tissues. J. Biol. Chem. 286, 5166–5174 (2011).
MacFarlane, A. J. et al. Cytoplasmic serine hydroxymethyltransferase regulates the metabolic partitioning of methylenetetrahydrofolate but is not essential in mice. J. Biol. Chem. 283, 25846–25853 (2008).
Mejia, N. R. & MacKenzie, R. E. NAD-dependent methylenetetrahydrofolate dehydrogenase-methenyltetrahydrofolate cyclohydrolase in transformed cells is a mitochondrial enzyme. Biochem. Biophys. Res. Commun. 155, 1–6 (1988).
Krupenko, S. A. & Oleinik, N. V. 10-formyltetrahydrofolate dehydrogenase, one of the major folate enzymes, is down-regulated in tumor tissues and possesses suppressor effects on cancer cells. Cell Growth Differ. 13, 227–236 (2002).
Anguera, M. C. et al. Regulation of folate-mediated one-carbon metabolism by 10-formyltetrahydrofolate dehydrogenase. J. Biol. Chem. 281, 18335–18342 (2006).
Krupenko, N. I. et al. ALDH1L2 is the mitochondrial homolog of 10-formyltetrahydrofolate dehydrogenase. J. Biol. Chem. 285, 23056–23063 (2010).
Sena, L. A. & Chandel, N. S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 48, 158–167 (2012).
Liou, G. Y. & Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 44, 479–496 (2010).
Piskounova, E. et al. Oxidative stress inhibits distant metastasis by human melanoma cells. Nature 527, 186–191 (2015).
Porporato, P. E. et al. A mitochondrial switch promotes tumor metastasis. Cell Rep. 8, 754–766 (2014).
Porporato, P. E., Payen, V. L., Baselet, B. & Sonveaux, P. Metabolic changes associated with tumor metastasis, part 2: mitochondria, lipid and amino acid metabolism. Cell. Mol. Life Sci. 73, 1349–1363 (2015).
Subramani, R. et al. Nimbolide inhibits pancreatic cancer growth and metastasis through ROS-mediated apoptosis and inhibition of epithelial-to-mesenchymal transition. Sci. Rep. 6, 19819 (2016).
Cheung, E. C. et al. Opposing effects of TIGAR- and RAC1-derived ROS on Wnt-driven proliferation in the mouse intestine. Genes Dev. 30, 52–63 (2016).
Gorrini, C., Harris, I. S. & Mak, T. W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 12, 931–947 (2013).
Jiang, L. et al. Reductive carboxylation supports redox homeostasis during anchorage-independent growth. Nature 532, 255–258 (2016).
Ye, J. et al. Serine catabolism regulates mitochondrial redox control during hypoxia. Cancer Discov. 4, 1406–1417 (2014). This paper describes a physiological role of serine in controlling mitochondrial ROS levels under conditions of hypoxia.
Ost, M. et al. Muscle mitohormesis promotes cellular survival via serine/glycine pathway flux. FASEB J. 29, 1314–1328 (2015).
Mehrmohamadi, M., Liu, X., Shestov, A. A. & Locasale, J. W. Characterization of the usage of the serine metabolic network in human cancer. Cell Rep. 9, 1507–1519 (2014).
Struck, A. W., Thompson, M. L., Wong, L. S. & Micklefield, J. S-Adenosyl-methionine-dependent methyltransferases: highly versatile enzymes in biocatalysis, biosynthesis and other biotechnological applications. Chembiochem 13, 2642–2655 (2012).
Kinnaird, A., Zhao, S., Wellen, K. E. & Michelakis, E. D. Metabolic control of epigenetics in cancer. Nat. Rev. Cancer https://dx.doi.org/10.1038/nrc.2016.82 (2016).
Sunden, S. L., Renduchintala, M. S., Park, E. I., Miklasz, S. D. & Garrow, T. A. Betaine-homocysteine methyltransferase expression in porcine and human tissues and chromosomal localization of the human gene. Arch. Biochem. Biophys. 345, 171–174 (1997).
Scott, J. M. & Weir, D. G. The methyl folate trap. A physiological response in man to prevent methyl group deficiency in kwashiorkor (methionine deficiency) and an explanation for folic-acid induced exacerbation of subacute combined degeneration in pernicious anaemia. Lancet 2, 337–340 (1981).
Maddocks, O. D., Labuschagne, C. F., Adams, P. D. & Vousden, K. H. Serine metabolism supports the methionine cycle and DNA/RNA methylation through de novo ATP synthesis in cancer cells. Mol. Cell 61, 210–221 (2016).
Mentch, S. J. et al. Histone methylation dynamics and gene regulation occur through the sensing of one-carbon metabolism. Cell Metab. 22, 861–873 (2015).
Hardie, D. G. AMP-activated protein kinase: an energy sensor that regulates all aspects of cell function. Genes Dev. 25, 1895–1908 (2011).
Ballatori, N. et al. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem. 390, 191–214 (2009).
Hultberg, B. & Hultberg, M. High glutathione turnover in human cell lines revealed by acivicin inhibition of gamma-glutamyltranspeptidase and the effects of thiol-reactive metals during acivicin inhibition. Clin. Chim. Acta 349, 45–52 (2004).
DeNicola, G. M. et al. NRF2 regulates serine biosynthesis in non-small cell lung cancer. Nat. Genet. 47, 1475–1481 (2015). A link between oxidative stress and activation of the SSP through NRF2 and ATF4.
Gruning, N. M. et al. Pyruvate kinase triggers a metabolic feedback loop that controls redox metabolism in respiring cells. Cell Metab. 14, 415–427 (2011).
Anastasiou, D. et al. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 334, 1278–1283 (2011).
Snell, K. Enzymes of serine metabolism in normal, developing and neoplastic rat tissues. Adv. Enzyme Regul. 22, 325–400 (1984).
Snell, K., Natsumeda, Y., Eble, J. N., Glover, J. L. & Weber, G. Enzymic imbalance in serine metabolism in human colon carcinoma and rat sarcoma. Br. J. Cancer 57, 87–90 (1988).
Pollari, S. et al. Enhanced serine production by bone metastatic breast cancer cells stimulates osteoclastogenesis. Breast Cancer Res. Treat. 125, 421–430 (2011).
Possemato, R. et al. Functional genomics reveal that the serine synthesis pathway is essential in breast cancer. Nature 476, 346–350 (2011).
Mullarky, E., Mattaini, K. R., Vander Heiden, M. G., Cantley, L. C. & Locasale, J. W. PHGDH amplification and altered glucose metabolism in human melanoma. Pigment Cell Melanoma Res. 24, 1112–1115 (2011).
Locasale, J. W. et al. Phosphoglycerate dehydrogenase diverts glycolytic flux and contributes to oncogenesis. Nat. Genet. 43, 869–874 (2011). References 75–77 show that overexpression of SSP enzymes contributes to the proliferation of some cancers.
Antonov, A. et al. Bioinformatics analysis of the serine and glycine pathway in cancer cells. Oncotarget 5, 11004–11013 (2014).
Pacold, M. E. et al. A PHGDH inhibitor reveals coordination of serine synthesis and one-carbon unit fate. Nat. Chem. Biol. 12, 452–458 (2016).
Liu, J. et al. Phosphoglycerate dehydrogenase induces glioma cells proliferation and invasion by stabilizing forkhead box M1. J. Neurooncol. 111, 245–255 (2013).
Fan, J. et al. Human phosphoglycerate dehydrogenase produces the oncometabolite D-2-hydroxyglutarate. ACS Chem. Biol. 10, 510–516 (2015).
Yoon, S. et al. Clinical implication of serine metabolism-associated enzymes in colon cancer. Oncology 89, 351–359 (2015).
Gylfe, A. E. et al. Eleven candidate susceptibility genes for common familial colorectal cancer. PLoS Genet. 9, e1003876 (2013).
Peters, J. M., Shah, Y. M. & Gonzalez, F. J. The role of peroxisome proliferator-activated receptors in carcinogenesis and chemoprevention. Nat. Rev. Cancer 12, 181–195 (2012).
Ericsson, A., Turner, N., Hansson, G. I., Wallenius, K. & Oakes, N. D. Pharmacological PPARα activation markedly alters plasma turnover of the amino acids glycine, serine and arginine in the rat. PLoS ONE 9, e113328 (2014).
Nilsson, L. M. et al. Mouse genetics suggests cell-context dependency for Myc-regulated metabolic enzymes during tumorigenesis. PLoS Genet. 8, e1002573 (2012).
Field, M. S. et al. Nuclear enrichment of folate cofactors and methylenetetrahydrofolate dehydrogenase 1 (MTHFD1) protect de novo thymidylate biosynthesis during folate deficiency. J. Biol. Chem. 289, 29642–29650 (2014).
MacFarlane, A. J., McEntee, M. F. & Stover, P. J. Azoxymethane-induced colon carcinogenesis in mice occurs independently of de novo thymidylate synthesis capacity. J. Nutr. 144, 419–424 (2014).
Macfarlane, A. J., Perry, C. A., McEntee, M. F., Lin, D. M. & Stover, P. J. Shmt1 heterozygosity impairs folate-dependent thymidylate synthesis capacity and modifies risk of Apc(min)-mediated intestinal cancer risk. Cancer Res. 71, 2098–2107 (2011).
Anderson, D. D., Woeller, C. F., Chiang, E. P., Shane, B. & Stover, P. J. Serine hydroxymethyltransferase anchors de novo thymidylate synthesis pathway to nuclear lamina for DNA synthesis. J. Biol. Chem. 287, 7051–7062 (2012).
Field, M. S., Kamynina, E., Watkins, D., Rosenblatt, D. S. & Stover, P. J. Human mutations in methylenetetrahydrofolate dehydrogenase 1 impair nuclear de novo thymidylate biosynthesis. Proc. Natl Acad. Sci. USA 112, 400–405 (2015).
MacFarlane, A. J., Perry, C. A., McEntee, M. F., Lin, D. M. & Stover, P. J. Mthfd1 is a modifier of chemically induced intestinal carcinogenesis. Carcinogenesis 32, 427–433 (2011).
Lee, G. Y. et al. Comparative oncogenomics identifies PSMB4 and SHMT2 as potential cancer driver genes. Cancer Res. 74, 3114–3126 (2014).
Gustafsson Sheppard, N. et al. The folate-coupled enzyme MTHFD2 is a nuclear protein and promotes cell proliferation. Sci. Rep. 5, 15029 (2015).
Lehtinen, L. et al. High-throughput RNAi screening for novel modulators of vimentin expression identifies MTHFD2 as a regulator of breast cancer cell migration and invasion. Oncotarget 4, 48–63 (2013).
Valencia, T. et al. Metabolic reprogramming of stromal fibroblasts through p62-mTORC1 signaling promotes inflammation and tumorigenesis. Cancer Cell 26, 121–135 (2014). Inhibition of the SSP in tumour-associated fibroblasts is shown to increase ROS levels, leading to inflammation and enhanced tumour progression.
Bollig-Fischer, A., Dewey, T. G. & Ethier, S. P. Oncogene activation induces metabolic transformation resulting in insulin-independence in human breast cancer cells. PLoS ONE 6, e17959 (2011).
Jun, D. Y. et al. Positive regulation of promoter activity of human 3-phosphoglycerate dehydrogenase (PHGDH) gene is mediated by transcription factors Sp1 and NF-Y. Gene 414, 106–114 (2008).
Ding, J. et al. The histone H3 methyltransferase G9A epigenetically activates the serine-glycine synthesis pathway to sustain cancer cell survival and proliferation. Cell Metab. 18, 896–907 (2013).
Wu, K. C., Cui, J. Y. & Klaassen, C. D. Beneficial role of Nrf2 in regulating NADPH generation and consumption. Toxicol. Sci. 123, 590–600 (2011).
Mitsuishi, Y. et al. Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell 22, 66–79 (2012).
Zhao, E. et al. KDM4C and ATF4 cooperate in transcriptional control of amino acid metabolism. Cell Rep. 14, 506–519 (2016).
Berry, W. L. & Janknecht, R. KDM4/JMJD2 histone demethylases: epigenetic regulators in cancer cells. Cancer Res. 73, 2936–2942 (2013).
Riscal, R. et al. Chromatin-bound MDM2 regulates serine metabolism and redox homeostasis independently of p53. Mol. Cell 62, 890–902 (2016).
Moran, D. M. et al. KRAS mutation status is associated with enhanced dependency on folate metabolism pathways in non-small cell lung cancer cells. Mol. Cancer Ther. 13, 1611–1624 (2014).
Audet-Walsh, E. et al. The PGC-1α/ERRα axis represses one-carbon metabolism and promotes sensitivity to anti-folate therapy in breast cancer. Cell Rep. 14, 920–931 (2016).
Ben-Sahra, I., Hoxhaj, G., Ricoult, S. J., Asara, J. M. & Manning, B. D. mTORC1 induces purine synthesis through control of the mitochondrial tetrahydrofolate cycle. Science 351, 728–733 (2016). This study describes a mechanism by which mTOR signals to increase purine synthesis by activating mitochondrial folate metabolism.
Ma, L. et al. Control of nutrient stress-induced metabolic reprogramming by PKCζ in tumorigenesis. Cell 152, 599–611 (2013).
Ou, Y., Wang, S. J., Jiang, L., Zheng, B. & Gu, W. p53 protein-mediated regulation of phosphoglycerate dehydrogenase (PHGDH) is crucial for the apoptotic response upon serine starvation. J. Biol. Chem. 290, 457–466 (2015).
Amelio, I. et al. p73 regulates serine biosynthesis in cancer. Oncogene 33, 5039–5046 (2014).
Melino, G., De Laurenzi, V. & Vousden, K. H. p73: friend or foe in tumorigenesis. Nat. Rev. Cancer 2, 605–615 (2002).
Kruiswijk, F., Labuschagne, C. F. & Vousden, K. H. p53 in survival, death and metabolic health: a lifeguard with a licence to kill. Nat. Rev. Mol. Cell Biol. 16, 393–405 (2015).
Labuschagne, C. F., van den Broek, N. J., Mackay, G. M., Vousden, K. H. & Maddocks, O. D. Serine, but not glycine, supports one-carbon metabolism and proliferation of cancer cells. Cell Rep. 7, 1248–1258 (2014). An unexpected inability of tumour cells to use glycine to support the one-carbon cycle and purine production.
Zhang, W. C. et al. Glycine decarboxylase activity drives non-small cell lung cancer tumor-initiating cells and tumorigenesis. Cell 148, 259–272 (2012). A paper describing the importance of glycine cleavage for tumour development.
Kim, D. et al. SHMT2 drives glioma cell survival in ischaemia but imposes a dependence on glycine clearance. Nature 520, 363–367 (2015). A study reconciling the observations in references 113 and 114, showing the importance of glycine cleavage for glycine detoxification, rather than contributing to the one-carbon cycle.
Azize, N. A. et al. Mutation analysis of glycine decarboxylase, aminomethyltransferase and glycine cleavage system protein-H genes in 13 unrelated families with glycine encephalopathy. J. Hum. Genet. 59, 593–597 (2014).
Vousden, K. H. & Lu, X. Live or let die: the cell's response to p53. Nat. Rev. Cancer 2, 594–604 (2002).
Start, K. Treating phenylketonuria by a phenylalanine-free diet. Prof. Care Mother Child 8, 109–110 (1998).
Hayashi, S., Tanaka, T., Naito, J. & Suda, M. Dietary and hormonal regulation of serine synthesis in the rat. J. Biochem. 77, 207–219 (1975).
Gravel, S. P. et al. Serine deprivation enhances antineoplastic activity of biguanides. Cancer Res. 74, 7521–7533 (2014).
Mullarky, E. et al. Identification of a small molecule inhibitor of 3-phosphoglycerate dehydrogenase to target serine biosynthesis in cancers. Proc. Natl Acad. Sci. USA 113, 1778–1783 (2016).
Chen, J. et al. Phosphoglycerate dehydrogenase is dispensable for breast tumor maintenance and growth. Oncotarget 4, 2502–2511 (2013).
Rose, M. L. et al. Dietary glycine prevents the development of liver tumors caused by the peroxisome proliferator WY-14,643. Carcinogenesis 20, 2075–2081 (1999).
Rose, M. L., Madren, J., Bunzendahl, H. & Thurman, R. G. Dietary glycine inhibits the growth of B16 melanoma tumors in mice. Carcinogenesis 20, 793–798 (1999).
Locasale, J. W. Serine, glycine and one-carbon units: cancer metabolism in full circle. Nat. Rev. Cancer 13, 572–583 (2013).
Gonen, N. & Assaraf, Y. G. Antifolates in cancer therapy: structure, activity and mechanisms of drug resistance. Drug Resist. Updat. 15, 183–210 (2012).
Longley, D. B., Harkin, D. P. & Johnston, P. G. 5-Fluorouracil: mechanisms of action and clinical strategies. Nat. Rev. Cancer 3, 330–338 (2003).
Huennekens, F. M. The methotrexate story: a paradigm for development of cancer chemotherapeutic agents. Adv. Enzyme Regul. 34, 397–419 (1994).
Valik, D., Radina, M., Sterba, J. & Vojtesek, B. Homocysteine: exploring its potential as a pharmacodynamic biomarker of antifolate chemotherapy. Pharmacogenomics 5, 1151–1162 (2004).
Dervieux, T., Greenstein, N. & Kremer, J. Pharmacogenomic and metabolic biomarkers in the folate pathway and their association with methotrexate effects during dosage escalation in rheumatoid arthritis. Arthritis Rheum. 54, 3095–3103 (2006).
Chang, C. & Pearce, E. Emerging concepts of T cell metabolism as a target of immunotherapy. Nat. Immunol. 17, 364–368 (2016).
Sharma, P. & Allison, J. P. The future of immune checkpoint therapy. Science 348, 56–61 (2015).
Acknowledgements
We would like to thank O. Maddocks and C. Labuschagne for helpful comments on the manuscript. This work was supported by Cancer Research UK grant C596/A10419 and European Research Council Grant 322842-METABOp53. We apologize to authors of studies that have contributed to this field but that we have been unable to cite owing to space constraints.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
K.H.V. is on the Science Advisory Board of Raze Therapeutics.
Glossary
- Non-essential amino acids
-
(NEAAs). Amino acids that can be synthesized de novo by the cell, and therefore need not be provided through an exogenous source, such as diet. These are alanine, arginine, asparagine, aspartic acid, cysteine, glutamic acid, glutamine, glycine, proline, serine and tyrosine. However, several of these become conditionally essential under certain conditions.
- Gluconeogenesis
-
Metabolic pathway for the generation of glucose from non-carbohydrate sources such as pyruvate, lactate or amino acids — essentially the reverse of glycolysis.
- Auxotrophic
-
Dependence on a nutrient for growth.
- One-carbon metabolism
-
The pathways that allow for the transfer of single carbon units through folate intermediates for use in purine and thymidylate synthesis, NADP+/NADPH production and methylation reactions. Although serine and glycine are major sources of one-carbon units, several other one-carbon donors are used by the cell, including choline, which is converted into betaine to provide one-carbon units in the remethylation of homocysteine to methionine. The dimethylglycine produced through this reaction can act as a further one-carbon source for the mitochondrial folate cycle.
- Pentose phosphate pathway
-
(PPP). An alternative pathway of glucose metabolism that produces NADPH for reductive biosynthesis and ribose 5-phosphate for nucleotide synthesis.
- Oxidative phosphorylation
-
Mitochondrial metabolic process that involves the transfer of electrons from NADH or FADH2 through the electron transport chain to reduce molecular oxygen to water. The energy released is used to regenerate ATP from ADP.
- Reactive oxygen species
-
(ROS). Reactive molecules derived from oxygen that can modulate the activity of redox-sensitive proteins or cause damage to nucleic acids, proteins and lipids.
- Oncometabolite
-
A metabolite whose abnormal accumulation is associated with cancer development.
- Glutaminolysis
-
Metabolic pathway of glutamine metabolism, allowing the use of carbon and nitrogen from glutamine to provide energy and anabolic intermediates.
Rights and permissions
About this article

Cite this article
Yang, M., Vousden, K. Serine and one-carbon metabolism in cancer. Nat Rev Cancer 16, 650–662 (2016). https://doi.org/10.1038/nrc.2016.81
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrc.2016.81
This article is cited by
-
Metabolic network of the gut microbiota in inflammatory bowel disease
Inflammation and Regeneration (2024)
-
Inhibition of mitochondrial folate metabolism drives differentiation through mTORC1 mediated purine sensing
Nature Communications (2024)
-
SHMT2 promotes papillary thyroid cancer metastasis through epigenetic activation of AKT signaling
Cell Death & Disease (2024)
-
Dysregulated cellular metabolism in atherosclerosis: mediators and therapeutic opportunities
Nature Metabolism (2024)
-
Serine synthesis promotes bone degradation
Nature Metabolism (2024)
