
Key Points
-
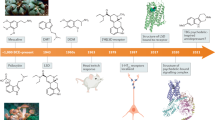
All antidepressant medications in use today are based on serendipitous discoveries, made more than 50 years ago, of the clinical efficacy of tricyclic and monoamine oxidase inhibitor antidepressants, both of which were later found to target the brain's serotonin or noradrenaline systems.
-
As only about half of all individuals with depression show full remission in response to these mechanisms, there has been intense interest in developing novel antidepressants with non-monoamine-based mechanisms. Although this work has led to impressive gains in understanding the molecular basis of adaptive and maladaptive responses to chronic stress, and in identifying numerous targets for the development of novel antidepressant treatments, it is striking that no non-monoamine-based antidepressant has yet been released onto the market.
-
We summarize the obstacles responsible for this lack of progress, including the difficult chemistry involved in most non-monoamine-based targets, limitations in animal models of depression, our still limited knowledge of the aetiology and pathophysiology of human depression, the heterogeneity and imprecise diagnosis of depression, and the exorbitant cost and risks of antidepressant clinical trials.
-
We go on to provide a progress report of some of the most promising current strategies aimed towards the development of non-monoamine-based antidepressants. Because a large subset of individuals with depression show hyperactivity of the hypothalamic–pituitary–adrenal (HPA) axis, the development of corticotropin-releasing factor (CRF) receptor antagonists has been a high priority. It is sobering, however, that despite decades of research, we still do not have a clear proof of concept demonstration of the efficacy of CRF antagonists in human depression. By contrast, there is increasing evidence for the possible antidepressant effects of glucocorticoid receptor antagonists.
-
By analogy with CRF antagonists, experience with substance P (or neurokinin, NK) receptor antagonists has been similarly frustrating. After an initial positive report almost a decade ago, more recent clinical evidence has been inconsistent, and the future of NK receptor antagonists in the treatment of depression remains uncertain.
-
More recent studies in animal models have shown putative roles for glutamate receptors, neurotrophic factors (such as brain-derived neurotrophic factor), the intracellular cyclic AMP pathway, hypothalamic feeding peptides, circadian genes, and many others in animal models of depression. These discoveries have defined new potential targets for antidepressant drug discovery, but real challenges remain in translating these to the clinic.
-
The identification of new targets such as these is cause for optimism. Nevertheless, past failures and frustrations in antidepressant drug development mean that a careful analysis of future advances is required.
Abstract
All available antidepressant medications are based on serendipitous discoveries of the clinical efficacy of two classes of antidepressants more than 50 years ago. These tricyclic and monoamine oxidase inhibitor antidepressants were subsequently found to promote serotonin or noradrenaline function in the brain. Newer agents are more specific but have the same core mechanisms of action in promoting these monoamine neurotransmitters. This is unfortunate, because only ∼50% of individuals with depression show full remission in response to these mechanisms. This review summarizes the obstacles that have hindered the development of non-monoamine-based antidepressants, and provides a progress report on some of the most promising current strategies.
Similar content being viewed by others

Main
Depression is a chronic, recurring and potentially life-threatening illness that affects up to 20% of the population across the globe1,2,3,4. It is one of the top ten causes of morbidity and mortality worldwide based on a survey by the World Health Organization. It is highly heritable, with roughly 40–50% of the risk for depression being genetic, although the specific genes that underlie this risk have not yet been identified. The remaining 50–60% of the non-genetic risk also remains poorly defined, with suggestions that early childhood trauma, emotional stress, physical illness, and even viral infections might be involved. Most experts agree that depression should be viewed as a syndrome, not a disease. Therefore, the highly variable compilation of symptoms that is used to define depression (Box 1), and the highly variable course of the illness and its response to various treatments, indicate that depression subsumes numerous disease states of distinct aetiology, and perhaps distinct pathophysiology. In fact, the lack of bona fide objective diagnostic tests for depression, beyond a compilation of symptoms, means that the diagnosis of the syndrome is quite variable, with no clear line distinguishing people who have mild clinical depression from those who are simply having a tough time in the course of normal life.
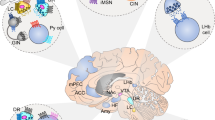
One key factor in the lack of objective diagnostic tests for depression is our still limited knowledge of the brain regions and neural circuits that are involved in the condition: if a biopsy were to be carried out in patients with depression, it is far from clear where the biopsy would be taken. Moreover, given the heterogeneity of the illness, different regions might well be involved in different individuals. Although the site of the pathology is unknown, there is growing knowledge of the brain regions that might mediate the diverse symptoms of depression1,2,3,4,5 (Box 2). The hippocampus and frontal regions of the cerebral cortex have received the most attention, particularly in animal studies of depression. These regions are expected to be particularly associated with cognitive abnormalities that are seen in many patients with depression. The amygdala, best studied for its role in establishing associations between aversive or rewarding stimuli and their associated environmental cues, has also been implicated. A role for the brain's reward pathways — for example, dopaminergic neurons in the ventral tegmental area and their target regions, in particular, the nucleus accumbens — has been proposed based on the prevalence of anhedonia and decreased motivation and energy levels in most individuals with depression. Similarly, abnormalities in appetite, sleep and circadian rhythms suggest the involvement of the hypothalamus as well. Human brain imaging studies and examination of postmortem brain tissue from people with depression support the contribution of these and several other brain regions to depression, but, so far, no clear consensus has evolved6,7,8,9.
In this review, we provide an overview of the mechanisms of action of currently available antidepressant treatments. As detailed below, all available antidepressants act via the monoamine neurotransmitters, serotonin or noradrenaline, and are based on serendipitous discoveries made in the 1950s. We then discuss the lack of success so far in developing antidepressants with non-monoamine-based mechanisms of action, and provide a progress report on some of the most promising strategies used in today's antidepressant drug discovery efforts.
Current antidepressant treatments
Despite the relative lack of knowledge of the aetiology and pathophysiology of depression, there are good treatments for it, with most patients showing significant improvement with optimal treatment. Mild depression responds to different forms of psychotherapy (Table 1). Mild and more severe forms of depression respond to a host of antidepressant medications, with a combination of medication and psychotherapy providing optimal treatment. electroconvulsive therapy (shock treatment) is one of most effective treatments for depression, but is usually reserved for the more severely ill due to the availability of numerous pharmacotherapies. The utility of other so-called somatic therapies is under investigation (Table 1).
Almost all of the available medications for depression are based on chance discoveries that were made more than half a century ago. Most of today's medications are based on the tricyclic antidepressants, which are believed to act by inhibiting the plasma membrane transporters for serotonin and/or noradrenaline1,2,3,10. These older medications provided a template for the development of newer classes of antidepressant, including the SSRIs (selective serotonin reuptake inhibitors), NRIs (noradrenaline reuptake inhibitors) and SNRIs (serotonin and noradrenaline reuptake inhibitors) (Table 1). However, as these newer medications have the same mechanism of action as the older tricyclics, their intrinsic efficacy and range of patients for whom treatment is successful remain the same. The older monoamine oxidase inhibitors, which reduce the enzymatic breakdown of serotonin and noradrenaline, are also still used today with great success.
Knowledge of the acute mechanisms of action of these drugs led to the general belief that all effective antidepressant medications act by increasing the activity of the brain's serotonergic or noradrenergic system. However, all of these medications must be given for at least several weeks for their antidepressant actions to become manifest. Despite several decades of research, and many interesting and promising leads, the changes that the drugs induce in the brain that underlie their therapeutic actions remain unclear.
Although today's treatments for depression are generally safe and effective, they are far from ideal. In addition to the need to administer the drugs for weeks or months to see clinical benefit, side effects are still a serious problem even with the newer medications. And, most importantly, fewer than 50% of all patients with depression show full remission with optimized treatment, including trials on numerous medications with and without concurrent psychotherapy. Therefore there is still a great need for faster acting, safer and more effective treatments for depression.
The search for novel antidepressants
Based largely on the acute pharmacological mechanisms of action of the older tricyclic and monoamine oxidase inhibitor medications, and the newer more selective serotonin and noradrenaline transporter inhibitors, the majority of antidepressant drug discovery efforts during the past few generations have focused on finding more selective serotonin or noradrenaline receptor agonists or antagonists, which might produce actions like those of the already available drugs, but more quickly and safely. Such efforts are still underway, with some promising leads. However, despite billions of dollars of research in both academia and industry, this approach has not yet succeeded in bringing any fundamentally new medications to the market. There are a handful of newer drugs known as atypical antidepressants, which have ascribed monoamine-based mechanisms, but there is only weak evidence that their purported mechanisms actually account for their clinical efficacy (Table 1).
At the same time, there has been an impressive accumulation of knowledge about non-monoamine systems that might contribute to the pathophysiology of depression in animal models, and some human evidence is also available1,2,3,4,5,11. However, none of these discoveries has so far been translated into a new bona fide treatment for depression. There are several reasons for this. First, it is not known whether the animal models that have been used to accurately predict the antidepressant action of serotonin- and noradrenaline-acting drugs (Box 3) can detect antidepressants that act through non-monoamine-based mechanisms. This is partly due to the fact that we have no bona fide non-monoamine-based antidepressants that have been adequately validated in humans. This is, therefore, a catch-22 situation (that is, one that cannot be resolved as it involves mutually conflicting or dependent conditions). Second, antidepressant efficacy studies are extremely expensive (they involve chronic treatment of at least hundreds of patients) and are notoriously risky (large placebo responses cause many trials to fail). This increases the threshold for a pharmaceutical or biotechnology company to embark on a trial of any antidepressant, especially one with a non-monoamine-based (and therefore riskier) mechanism. Third, to increase their confidence level in a non-monoamine-based drug, many groups have looked for effects of such drugs on the serotonin and noradrenaline systems. According to this view, if it can be shown that a non-monoamine-based drug enhances, for example, serotonergic transmission in some brain region, this increases the cache of that drug. However, this is another catch-22, as it does not lead us to create drugs with truly novel mechanisms of action. Finally, profits from monoamine-based drugs (SSRIs and SNRIs) have been extremely high, and this has removed the financial incentive to take the risks involved in developing drugs with non-monoamine-based actions.
Nevertheless, with expiring patents for most of the newer agents looming, academic and industrial scientists are increasingly of the opinion that the field must move beyond today's mechanisms of antidepressant medications. Below, we discuss some of the best hopes for non-monoamine-based drugs for the treatment of depression. Given space limitations, this review is not comprehensive; rather, we highlight only some examples of current non-monoamine approaches to antidepressant drug discovery, with some additional (more preliminary) examples given in Box 4.
However, at the outset, we must acknowledge a major challenge. The lack of progress in identifying validated depression vulnerability genes in humans, and lack of knowledge of specific environmental factors that interact with such depression genes to cause the illness, means that, at present, there is no perfect animal model for studies of depression or antidepressant action. This is particularly important, because antidepressants do not elevate mood in healthy humans. The absence of perfect animal models has forced the field to focus on available paradigms, most of which involve exposure of healthy animals (which do not have the genes that predispose certain humans to depression) to various forms of acute or chronic stress (Box 3). However, the relationship between stress and depression is controversial: it is far from certain that exposure to stress per se can induce depression in most healthy humans. Consequently, the clinical relevance of actions of putative antidepressants on stress-induced behavioural abnormalities in animal models remains unproven. Another complicating factor is the lack of clear distinction between depression and anxiety in both humans and animal models. Therefore, some depressed patients show strong symptoms of anxiety, whereas others are emotionally blunted (Table 1), and, as detailed below, some putative antidepressants are equally active in animal anxiety and depression models. Finally, it is ironic that the search for new targets for antidepressants has typically involved searching for proteins and genes that are altered in these models by stress, and showing reciprocal regulation by reuptake inhibitor antidepressants. Therefore, we have another catch-22: the search for non-monoamine-based antidepressants has often relied on the actions of monoamine-based drugs. The buyer beware!
CRF and glucocorticoids
Glucocorticoid release is controlled by the hypothalamic–pituitary–adrenal (HPA) axis. Corticotropin-releasing factor (CRF) released by the paraventricular nucleus of the hypothalamus stimulates the release of corticotropin (ACTH) from the anterior pituitary, which, in turn, stimulates glucocorticoid secretion from the adrenal cortex (Fig. 1). The HPA axis is an essential component of an individual's capacity to cope with stress. Excessive stimulation of the axis has been implicated in depression. Hyperactivity of the HPA axis is observed in the majority of patients with depression, as manifested by increased expression of CRF in the hypothalamus, increased levels of CRF in the cerebrospinal fluid (CSF), and reduced feedback inhibition of the axis by CRF and glucocorticoids3,12,13,14,15,16. Although the molecular basis of these derangements in the HPA axis remains unknown, the results of numerous clinical studies suggest that normalization of the axis might be a necessary step for stable remission of depressive symptoms. In animal models, hypercortisolaemia can potentiate excitotoxicity of hippocampal pyramidal neurons — as evidenced by dendritic atrophy and spine loss, and possibly cell death — as well as inhibit the birth of new granule cell neurons in the hippocampal dentate gyrus, and many of these changes can be prevented by antidepressant treatment11,12,15,17. Excessive glucocorticoids could, therefore, be a causative factor for the small reductions in hippocampal volume that have been reported in patients with depression or post-traumatic stress disorder, although this finding remains controversial (see Ref. 1).

Corticotropin-releasing factor (CRF) from the paraventricular nucleus (PVN) of the hypothalamus is released into the hypophyseal portal system and triggers the release of corticotropin (ACTH) from the anterior pituitary via stimulation of CRF1 receptors. ACTH, in turn, stimulates the secretion of glucocorticoid hormones (cortisol in humans or corticosterone in rodents) from the adrenal cortex. Increased glucocorticoid levels suppress hypothalamic CRF expression via negative feedback through hippocampal and hypothalamic glucocorticoid receptors. The neurotransmitter action of CRF on CRF1 receptors throughout the limbic system mediates anxiogenic effects of stress. By contrast, its neurotransmitter action on CRF2 receptors in more discrete regions of the brain might reduce anxiety-like behaviour in a delayed fashion. Amy, amygdala; BNST, bed nucleus of the stria terminalis; DR, dorsal raphe; HPA, hypothalamic–pituitary–adrenal; LDT, laterodorsal tegmental nucleus; LH, lateral hypothalamus; Pit, pituitary; Thal, thalamus.
CRF also serves as a neurotransmitter in several brain areas outside the hypothalamus — in particular, the central nucleus of the amygdala and bed nucleus of the stria terminalis (BNST) (Fig. 1). The amygdala neurons send wide projections to the forebrain and brainstem, and have a crucial role in negative emotional memory (for example, as measured by fear conditioning). The amygdala and BNST are implicated in the generation of anxiety-like behaviour18,19. Elevated levels of CRF have been found in some projection areas of these regions (for example, the locus coeruleus) in patients with depression3,7. An impressive literature has directed intense interest in the CRF and glucocorticoid systems as targets for the development of novel antidepressants.
CRF antagonists. Overexpression of CRF in transgenic mice, or CRF administration into the CNS, causes several depression-like symptoms, including hypercortisolaemia, decreased appetite and weight loss, and decreased sexual behaviour3,14,16,20,21,22. These conditions also increase arousal and induce anxiety-like behaviours. These various symptoms are presumably mediated through increased CRF function both in the HPA axis and in the amygdala, BNST and related circuits. Physiological actions of CRF are mediated through two types of receptor, CRF1 and CRF2, both of which are coupled to the Gs subunit of G proteins — the subunit that stimulates adenylyl cyclase to increase cyclic AMP (cAMP) synthesis. CRF1 is the predominant subtype: these receptors are enriched in the pituitary, where they regulate the HPA axis, and are also highly expressed throughout limbic brain regions, where their selective deletion attenuates behavioural responses to stress3,16,20,21,22. These data supported a massive effort to develop CRF1 antagonists as anxiolytic and antidepressant medications. Such compounds dramatically reduce anxiety-like behaviour and fear conditioning in rodents3,16,23, and also antagonize a range of depression-like symptoms seen during withdrawal from several drugs of abuse24. However, CRF1 antagonists have not shown consistent activity in standard antidepressant screens (for more information, see Ref. 23), which raises questions, stated earlier, about the relevance of rodent stress models to human depression. One open label clinical trial found that a non-peptidic CRF1 antagonist reduces depression and anxiety scores in patients with depression, without interference of the HPA axis25. However, so far, no well-controlled study has verified these findings. Unfortunately, pharmacokinetic and hepatotoxicity issues have led to the discontinuation of this and numerous other CRF1 antagonists26, which is an all too common occurrence for drugs aimed at neuropeptide receptors. The failure to obtain clear proof of concept of the CRF1 antagonist mechanism as either anxiolytic or antidepressant in humans, despite decades of research, is a major disappointment and frustration in the field.
CRF2 shows more restricted expression in the brain, and their role in regulating complex behaviour is still under investigation. CRF2-knockout mice show usual anxiety-like behaviour, but CRF2 antagonists show anxiolytic properties in animal models and some, but not all, also show significant efficacy in the learned helplessness and chronic mild stress depression paradigms20,22,23. Recent results indicate that the endogenous ligands for CRF2, in addition to CRF, may be the urocortin peptides, which promote adaptive responses to stress16,22. There remains considerable interest in the clinical development of CRF2 antagonists, particularly as they are less likely than CRF1 antagonists to cause side effects via the HPA axis.
Vasopressin receptor antagonists. The neuropeptide vasopressin, which is synthesized in the paraventricular and supraoptic hypothalamic nuclei, is well known for its role in fluid metabolism. It also regulates the HPA axis: stress stimulates the release of vasopressin, which then potentiates the effects of CRF on ACTH release. Vasopressin is also found outside the hypothalamus, notably in the amygdala and BNST, and is believed to exert effects throughout the limbic system through activation of vasopressin V1a and V1b receptors. Vasopressin levels are reportedly increased in some patients with depression and might contribute to HPA axis abnormalities observed in these individuals. Furthermore, in postmortem studies, SSRI treatment has been reported to normalize vasopressin levels27. Non-peptide V1b antagonists show antidepressant-like effects in rodents, partly through amygdala-dependent mechanisms28. This is in contrast to V1b-knockout mice, which show normal stress responses29,30. Vasopressin antagonists have yet to be evaluated in humans.
Glucocorticoids: agonists or antagonists? Glucocorticoids diffuse passively through cellular membranes and bind to intracellular glucocorticoid receptors (GR), causing their translocation into the nucleus3,16. In the nucleus, these ligand-activated transcription factors bind to specific DNA response elements, or to other transcription factors, and alter gene expression. Glucocorticoids also cause rapid effects at the neuronal plasma membrane through distinct proteins that remain incompletely characterized. In the brain, glucocorticoid-regulated genes affect many aspects of neuronal function, including metabolism, neuronal connections, and synaptic transmission. Glucocorticoids also promote the termination of stress reactions through complex feedback loops, mediated in part through the hippocampus and paraventricular nucleus, ultimately leading to the repression of target genes implicated in stress responses, such as CRF. Interestingly, glucocorticoids exert stimulatory effects on CRF expression in other circuits, for example, the amygdala and BNST, which further highlights the complexity of these systems16. Although most of the transcriptional effects of glucocorticoids are mediated through the GR2 receptor, these hormones also act at GR1 (mineralocorticoid receptor), which contributes to HPA axis physiology and stress responses as well15,16.
As mentioned earlier, insufficient feedback suppression of the HPA axis by CRF and glucocorticoids is seen in a large subset of patients with depression. Recently, this neuroendocrine abnormality was reproduced in adult mice with selective deletion of GR2 in the forebrain31. Interestingly, this mutation also resulted in a robust depression-like phenotype, and many of these abnormalities were corrected by chronic treatment with tricyclic antidepressants. Conversely, transgenic mice overexpressing GR2 in the forebrain are more sensitive to the acute effects of antidepressants32. These findings raise the possibility that enhanced GR activity in the forebrain might be antidepressant. Most antidepressant treatments can restore efficient negative feedback of the HPA axis, and increase the expression of GR in forebrain regions such as the hippocampus3,14,16. Some patients with depression carry a polymorphism, or genetic variant, in the FKBP5 gene (which encodes a co-chaperone of heat-shock protein 90 (HSP90)) that results in higher affinity of GR for cortisol33. These individuals reportedly respond to antidepressants much faster than those without this mutation.
These findings are paradoxical, given the evidence, cited above, that hypercortisolaemia might contribute to the pathophysiology of depression3,14,15,16, but the two sets of results could be reconciled as follows. Deficient inhibitory feedback of the HPA axis might result from excessive activation of GR in the hippocampus, and subsequent damage to this region12,15. Recently, viral vectors have been used to deliver into the hippocampus chimaeric GR that combined the ligand-binding domain of GR with the DNA-binding domain of oestrogen receptors, thereby converting the glucocorticoid signal into an oestrogen-like effect34,35. The expression of the chimaeric receptor potently reduced hippocampal damage and rendered excess glucocorticoids protective rather than destructive. The behavioural effects of such genetically altered GR have not yet been reported in animal models of depression.
There is increasing clinical evidence to suggest that depressive symptoms in patients with psychotic depression or cushing syndrome might be rapidly ameliorated by GR antagonists3,16,36. The GR antagonist mifepristone (which is also a progesterone receptor antagonist and is used clinically to induce chemical abortion of early pregnancy) is currently in Phase III clinical trials for psychotic major depression and might be the first non-monoaminergic-based antidepressant on the market37. Its use is also associated with alterations of the HPA axis36. The glucocorticoid synthesis inhibitor, metyrapone, also shows some promise in treating depression when added to a standard antidepressant38.
The neurokinin system
Substance P, a member of the tachykinin neuropeptide family, is the preferred endogenous agonist for neurokinin 1 (NK1) receptors, which are coupled to the Gq subunit of G proteins — the subunit that can stimulate phospholipase C (PLC). Substance P is the most abundant tachykinin in the CNS, where it has been studied primarily for its role as a central mediator of pain, an indication for which non-peptidic NK1 receptor antagonists were initially developed39. The rationale for considering NK1 receptor antagonists in depression was based on the expression of substance P and NK1 receptors in fear- and anxiety-related circuits, the release of substance P in animals in response to fearful stimuli, and the strong co-localization of substance P with serotonin and noradrenaline or their receptors in the human brain39,40,41. Reciprocally, local application of substance P agonists was shown to induce a range of neural, behavioural and cardiovascular changes characteristic of defensive responses, including increased firing of the locus coeruleus, place aversion, distress vocalizations, escape behaviour and cardiovascular activation. Moreover, some effects of stress can be blocked by systemic administration of NK1 receptor antagonists. These effects have since been confirmed by the anxiolytic- and antidepressant-like phenotype of substance P- and NK1 receptor-knockout mice40,41.
In 1998, Kramer et al. published the first evidence that chronic treatment with a non-peptidic NK1 receptor antagonist might be antidepressant in humans42. This report was greeted with great enthusiasm, but, although its results were replicated in some studies, replication failed in others, such that the validity of NK1 receptor antagonism as an effective antidepressant strategy remains uncertain. Indeed, several pharmaceutical companies have discontinued their NK1 receptor antagonist programmes in yet another big disappointment for the field37,39.
Although NK1 receptor antagonists were initially claimed to act through a completely novel mechanism of action, subsequent studies have suggested that their therapeutic action, if any, could be secondary to changes in monoaminergic systems. NK1 receptor antagonists have a delayed onset of action similar to that of monoamine-based antidepressants, and their chronic administration causes increased firing of serotonergic neurons — a change also observed in NK1 receptor-knockout mice42,43. In addition, genetic or pharmacological blockade of NK1 receptors induces hippocampal neurogenesis and some of the same long-term effects in the brain as do bona fide antidepressants on cell signalling proteins, such as induction of brain-derived neurotrophic factor (BDNF)43,44,45,46,47,48,49. These results raise the possibility that NK1 receptor antagonists could conceivably be used as augmentation agents in combination with a traditional antidepressant44.
BDNF and other neurotrophic mechanisms
The neurotrophic hypothesis of depression and antidepressant action was originally based on findings in rodents that acute or chronic stress decreases expression of BDNF in the hippocampus and that diverse classes of antidepressant treatment produce the opposite effect and prevent the actions of stress11,50 (Fig. 3). These observations led to the suggestion (still unproven) that perhaps such changes in BDNF could in part mediate the structural damage and reduced neurogenesis seen in the hippocampus after stress and the prevention of these effects by antidepressant treatments (see above). Importantly, on autopsy, reduced BDNF levels in the hippocampus have been reported in some patients with depression — an abnormality not seen in patients treated with antidepressants51.

a | Shows a normal hippocampal pyramidal neuron and its innervation by glutamatergic, monoaminergic and other types of neuron. Its regulation by brain-derived neurotrophic factor (BDNF), which is derived from the hippocampus or other brain areas, is also shown. b | Severe stress causes several changes in these neurons, including a reduction in their dendritic arborization, and a reduction in BDNF expression (which could be one of the factors mediating the dendritic effects). The reduction in BDNF is mediated partly by excessive glucocorticoids, which could interfere with the normal transcriptional mechanisms (for example, through cyclic AMP-responsive-element-binding protein, CREB) that control BDNF expression. c | Antidepressants produce the opposite effects to those seen in b: they increase dendritic arborization and BDNF expression of these hippocampal neurons. The latter effect appears to be mediated at least in part by activation of CREB. By these actions, antidepressants might reverse and prevent the effects of stress on the hippocampus, and ameliorate some symptoms of depression. Modified, with permission, from Ref. 2 © (2002) Cell Press.
Together, these data support the possibility that drugs that activate BDNF signalling in the hippocampus might be antidepressant. Direct evidence for this hypothesis comes from experiments in which injection of BDNF into the rodent hippocampus exerts antidepressant-like effects in the forced swim and learned helplessness tests52. Conversely, inducible knockout of BDNF from the hippocampus and other forebrain regions prevents the antidepressant effects of reuptake inhibitor antidepressants in these paradigms53.
Although a great deal of work remains to be done to validate this hypothesis, the main challenge from a drug discovery point of view is that BDNF is not an easy drug target. It is a small protein of 14 kDa, which binds to its TrkB tyrosine kinase receptor as a dimer. Accordingly, it is difficult to develop small molecule agonists of TrkB. On the basis of studies in cell culture, it is known that BDNF activation of TrkB leads to diverse physiological effects by regulating a complex cascade of post-receptor pathways, which involve Ras–Raf–ERK (extracellular-signal regulated kinase), phosphatidylinositol 3-kinase (PI3K)–Akt (v-akt murine thymoma viral oncogene homologue) and PLCγ. In theory, this raises the possibility of targeting numerous proteins for antidepressant development, however, several obstacles remain. First, we do not yet know which of these pathways are most crucial for the antidepressant actions of BDNF in animal models; second, most of these signalling proteins are broadly expressed throughout the brain and peripheral tissues, which heightens concerns about toxicity of any drug directed against them; and third, the lack of availability of small molecule agonists for most of these signalling proteins means that their potential antidepressant activity cannot easily be assessed (another catch-22). One potential strategy to overcome the last two obstacles is to target proteins in BDNF signalling cascades that are enriched in particular brain circuits implicated in depression.
Another complication is that, although BDNF might exert antidepressant-like effects at the level of the hippocampus, its actions might be different, or even opposite, in other neural circuits. The best example is the ventral tegmental area–nucleus accumbens dopaminergic reward circuit, in which chronic stress increases BDNF expression, local BDNF infusion exerts a prodepression-like effect in the forced swim test, and blockade of BDNF function exerts an antidepressant-like effect54,55. A more recent study found a similar antidepressant-like effect on viral-mediated local knockout of BDNF from the ventral tegmental area in a social defeat model55. These findings raise caution about the goal of developing an antidepressant based on BDNF, as a drug that promotes BDNF function might produce competing effects in different brain regions. This again emphasizes the approach, mentioned above, of targeting BDNF signalling proteins that show more restricted patterns of expression in the brain.
In addition to BDNF, other neurotrophic factors also warrant consideration as potential leads for antidepressant development11. A recent DNA microarray study of the human hippocampus found that several genes in the fibroblast growth factor (FGF) family — FGF and some of its receptors — are downregulated in the hippocampus of patients with depression56. This is interesting in light of the knowledge that FGF seems to be an important endogenous regulator of neurogenesis in the adult rat hippocampus. Still other neurotrophic factors are known to be regulated in the hippocampus by stress and antidepressant treatments, which are currently being evaluated in depression models11,57.
Studies of neurotrophic mechanisms in depression and antidepressant action have provided important heuristic models for the field. However, it may be difficult to translate these discoveries into new treatment approaches for depression due to the complexity of neurotrophic factors and their receptors and post-receptor signalling cascades.
Phosphodiesterase inhibitors
Phosphodiesterases (PDEs) catalyse the degradation of cAMP and cGMP. The potential antidepressant activity of phosphodiesterase inhibitors dates back decades to the idea that these drugs would be expected to promote the actions of noradrenaline at β-adrenergic receptors, which, at the time, were proposed to partly mediate antidepressant responses. Indeed, there were early indications that rolipram, a non-selective PDE4 inhibitor might be antidepressant in small clinical trials (for more information, see Refs 11, 50). These early trials failed because rolipram and related PDE4 inhibitors induced intense nausea and vomiting.
Renewed interest in PDE4 inhibitors as antidepressants has come from the finding that they induce BDNF expression in the hippocampus11,50. This effect seems to be mediated by activation of the cAMP pathway, which leads to the activation of the transcription factor cAMP-responsive-element (CRE)-binding protein (CREB) and to the direct induction of the Bdnf gene via a CRE site in its promoter (Fig. 3). Induction of CREB itself in the hippocampus exerts an antidepressant-like effect in the forced swim test58. Therefore, PDE4 inhibitors might provide an indirect way to promote CREB and BDNF function, and exert an antidepressant effect. Meanwhile, there is intense interest in PDE4 inhibitors as cognitive enhancers, a possibility that is also based on the role of CREB in the hippocampus — in this case in mediating important forms of learning and memory59,60. The main challenge, however, remains side effects: is it possible to inhibit PDE4 in the hippocampus and exert antidepressant effects and cognitive enhancement without inhibiting PDE4 in brainstem regions, which causes nausea and vomiting?
A second major challenge is that inhibition of phosphodiesterase isoforms might not be antidepressant or enhance cognition in all brain regions. There is growing evidence that stimulation of the cAMP pathway and CREB in the nucleus accumbens might be prodepressant. Therefore, mechanisms to oppose, rather than to enhance, activity of this pathway might be more suitable for antidepressant drug discovery efforts61 (Fig. 2). Similarly, stimulation of the cAMP pathway in frontal cortical regions can inhibit cognitive function in aged animals62, which again highlights potential problems of targeting phosphodiesterase isoforms that are widely expressed in the brain. On the positive side, there are four subtypes of PDE4, PDE4A–D, each of which is encoded by a different gene, with multiple splice variants of each subtype11. It is conceivable that a particular subtype enriched in the hippocampus could be targeted for antidepressant and cognition-enhancing effects, although this remains conjectural. In addition, there are many other phosphodiesterase isoforms, some of which show highly restricted patterns of expression in the brain. For example, PDE10A is highly enriched in the striatum. It, too, could potentially be targeted for antidepressant development. Moreover, there are many other families of signalling proteins that modulate G protein–adenylyl cyclase activity, such as regulators of G protein signalling (RGS) proteins, subtypes of which show restricted expression patterns in the brain. These proteins also represent potential drug targets1,2.

The figure shows a simplified hypothetical scheme, whereby the activation of CREB (cyclic AMP (cAMP)-responsive-element-binding protein) by stress mediates the induction of dynorphin (DYN) in the nucleus accumbens (NAc), which then contributes to anhedonia-like symptoms. CREB is activated by D1 dopamine receptors (through activation of the cAMP pathway) or by Ca2+- or tyrosine receptor kinase B (TrkB)-regulated signal transduction pathways, leading to increased expression of dynorphin. Dynorphin feeds back onto κ opioid receptors located on the terminals and cell bodies/dendrites of ventral tegmental area (VTA) dopamine (DA) neurons. Stimulation of these κ receptors inhibits the VTA neurons, which leads to anhedonia-related symptoms. Therefore, antagonists of κ receptors might block the consequences of CREB-induced increases in dynorphin activity, and exert antidepressant effects in some individuals. GABA, γ-aminobutyric acid.
Glutamate acting drugs
The link between glutamatergic neurotransmission and the pathophysiology of depression has been increasingly demonstrated since the 1950s, when the mood elevating properties of anti-infectious agents with some NMDA (N-methyl-D-aspartate) glutamate receptor antagonist activity (for example, D-cycloserine and amantadine) were first reported63,64. A rapid antidepressant effect of a single intravenous injection of ketamine, a dissociative anaesthetic and NMDA receptor antagonist, was subsequently shown in a placebo-controlled trial. The application of ketamine and related drugs as antidepressants is obviously limited by their severe psychotomimetic action. However, clinical trials are now assessing the antidepressant potential of the weaker NMDA receptor antagonist memantine, and the glutamate release inhibitor riluzole, two FDA (Food and Drug Administration, USA) approved compounds developed for cognitive enhancement and neuroprotection, respectively. Although clinical evidence supporting the antidepressant efficacy of NMDA receptor antagonists is still relatively weak, preclinical research increasingly suggests that reduced NMDA receptor function is antidepressant-like in several animal models and prevents stress-induced alterations in hippocampal neuronal morphology, and that chronic treatment with bona fide antidepressants downregulates NMDA receptors or reduces glutamate release through presynaptic mechanisms15,63,64. A recent report indicated that deletion of a novel NMDA receptor subunit causes an anxiolytic- and antidepressant-like profile65.
It has been reported that activation of AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid) glutamate receptors increases BDNF expression, and rapidly stimulates neurogenesis and neuronal sprouting, in the hippocampus11. Based on these observations, another strategy has been the evaluation of AMPA receptor potentiators in models of depression63,64,66. Positive allosteric modulators, which avoid the rapid desensitization of AMPA receptors seen with full agonists, were reported to have similar activity to tricyclics and SSRIs in the forced swim and tail suspension tests. Interestingly, AMPA receptor potentiators were also active in reducing rat submissive behaviour (a behavioural model that responds selectively to chronic antidepressant treatment) with a shorter onset of action than an SSRI. There are also some indications that monoamine-based antidepressants promote AMPA receptor function.
Given the dominant role of ionotropic glutamate receptors in synaptic activity and plasticity throughout the brain, including cognition-, emotion- and reward-related circuits, it is not surprising that agents that affect these receptors could exert antidepressant activity. It remains to be seen whether such drugs could have the selectivity and safety required. One proposed strategy would be to target any of several metabotropic (or G-protein-coupled) glutamate receptors, which seem to differentially modulate the activity of the ionotropic receptors and might thereby mediate safer and more selective effects67.
Hypothalamic feeding peptides
During the past decade, there have been explosive advances in understanding hypothalamic peptides that regulate feeding behaviour. Recent work has begun to draw connections between these hypothalamic feeding peptides and depression. Of particular note is melanin-concentrating hormone (MCH), a major orexigenic (pro-appetite) peptide expressed in a subset of lateral hypothalamic neurons. The MCH1 receptor, the only subtype expressed in rodents, is coupled to the inhibitory subunit of G proteins (Gi), and shows remarkable enrichment in the nucleus accumbens68. Direct administration of MCH into this region stimulates feeding behaviour, whereas blockade of the MCH1 receptor decreases feeding69. Intracerebroventricular and intrahypothalamic MCH administration have similar effects. Moreover, several MCH1 receptor antagonists — including non-peptidic small molecule antagonists — administered systemically or directly into the nucleus accumbens exert antidepressant-like effects in the forced swim test69,70. A similar antidepressant-like phenotype is observed in mice lacking MCH or the MCH1 receptor, whereas a prodepressant-like phenotype is seen in MCH-overexpressing animals69,71. Taken together, these data provide a strong case that MCH antagonists, by disrupting MCH signalling to the nucleus accumbens, might provide a novel mechanism for antidepressant medications. These drugs would also reduce weight, which could be particularly useful in the subset of patients with depression who show weight gain. Evaluating these agents in humans is now the main obstacle.
Several other hypothalamic feeding peptides also deserve attention in the depression field. These include orexigenic peptides, such as orexin (hypocretin), neuropeptide Y (NPY), and agouti-related peptide (ARP), as well as anorexigenic peptides, such as melanocortin (α-MSH), cocaine- and amphetamine-regulated transcript (CART), and CRF. Many of these peptides have been shown to not only regulate feeding, but to alter reward mechanisms, which suggests that they could have possible effects on anhedonia-related symptoms4,5,72. Some of the peptides, such as CRF (see above) and NPY4, also deserve attention as antidepressant targets, because they are expressed — well beyond the hypothalamus — in limbic brain circuits, where they have been implicated in depression- and anxiety-like behaviours. Interestingly, these feeding peptide systems could produce very different effects in different subtypes of depression. For example, individuals whose depression is characterized by reduced activity and weight gain might respond to different agents from depressed individuals who show increased activity, anxiety and weight loss.
Circadian gene products
Abnormal circadian rhythms have long been described in depression and other mood disorders (Box 1). Many patients with depression report their most serious symptoms in the morning with some improvement as the day progresses. This might represent an exaggeration of the diurnal fluctuations in mood, motivation, energy level and responses to rewarding stimuli that are commonly seen in the healthy population. The molecular basis for these rhythms seen under normal and pathological conditions is poorly understood.
Most research on circadian rhythms has focused on the suprachiasmatic nucleus (SCN) of the hypothalamus, which is considered the master circadian pacemaker of the brain73,74. Here, circadian rhythms are generated at the molecular level by clock (Clk, a Pas–domain-containing transcription factor), which dimerizes with Bmal (another transcription factor); and the dimer induces the expression of the genes Per (period) and Cry (cryptochrome), which, in turn, feed back to repress Clk–Bmal activity. In addition, Clk–Bmal, Per and Cry regulate the expression of many other genes, which presumably drive the many circadian variations in cell function. This molecular cycle in the SCN is entrained by light and seems to be essential for matching circadian rhythms with the light–dark cycle. However, more recent research has indicated that control of circadian rhythms is far more complicated than this simple model. Clk, Bmal, Per and Cry, as well as several related genes, are broadly expressed throughout the brain, including in limbic regions implicated in mood regulation, although little is known about their function outside the SCN.
Recent work has established that circadian genes regulate brain reward. For example, cocaine reward is markedly enhanced in mice that lack Clk, and this abnormality is associated with a dramatic increase in the activity of ventral tegmental area dopamine neurons75. Clk expression is regulated in the striatum and hippocampus by cocaine and antidepressants, and the results of preliminary studies suggest that Clk mutant mice show less depression-like behaviour in the forced swim test, as well as reduced brain stimulation reward thresholds, which indicates an elevated affective state76,77. Together, these studies are consistent with an important influence of Clk, at the level of the ventral tegmental area–nucleus accumbens pathway, and perhaps other circuits, in the regulation of mood, and suggest that abnormalities in circadian gene function could contribute to certain symptoms of depression. There has also been interest in a clock-like protein, known as NPAS2 (neuronal Pas domain protein 2), which dimerizes with Bmal to regulate the expression of Per, Cry and many other genes; this regulation, like that mediated by Clk, shows circadian rhythms78. Interestingly, NPAS2 is not expressed in the SCN, but is found at high levels in several limbic regions, particularly the nucleus accumbens. Npas2-knockout mice show increased anxiety-like behaviour and deficits in fear conditioning78. In addition, the mice show deficits in their ability to entrain to non-light stimuli, such as food79. It has been suggested that NPAS2 is a crucial mediator of circadian rhythms in an individual's emotional state through actions in the nucleus accumbens and other limbic regions. Glycogen synthase kinase 3β (GSK3β) is one of several kinases involved in regulation of circadian cycles through the phosphorylation of Per and other circadian gene products73,74. GSK3β also regulates many other biochemical pathways, for example, Wnt signalling through β-catenin. The kinase is one of many known acute targets of lithium and other mood-stabilizing agents, and could represent another potential connection between circadian rhythms and treatment of mood disorders80,81.
Taken together, the results of these early studies support the hypothesis that circadian genes might function abnormally in depression and other mood disorders. This work also suggests that drugs aimed at influencing particular target genes for these circadian transcription factors, which are expressed in distinct brain circuits, deserve attention as targets for possible new treatment agents for depression.
Future directions
Antidepressant drug discovery is at a crossroads. Available medications with monoamine-based mechanisms will be going off patent during the next decade, and proof of concept studies for some of the best neuropeptide and neuroendocrine targets (for example, CRF, substance P and glucocorticoid receptors), which are based largely on stress models, should at last be available within the next few years. At the same time, a host of fundamentally new targets has emerged as a result of more open-ended molecular and cellular approaches in concert with improving, albeit still imperfect, animal models of stress82,83. Progress with some of these targets (for example, BDNF) has been hampered by the difficult chemistry involved. Nevertheless, this research has suggested numerous biomarkers or endophenotypes for depression1,2,5,11 — for example, BDNF expression, hippocampal neurogenesis, neuronal morphology and CREB activity, to name just a few. However, it is difficult to measure any of these biomarkers or endophenotypes in living patients.
A considerable leap forward in the field will require identification of genes that confer risk for depression in humans, and understanding how specific types of environmental factor interact synergistically with genetic vulnerability. This will make it possible to develop more valid animal models of human depression. Important advances will also require the development of ever more penetrating brain imaging methodologies to enable the detection of molecular and cellular biomarkers in living patients. Such discoveries should at last make it possible to delineate bona fide subtypes of depression, which will probably show distinct aetiological and pathophysiological mechanisms.
Ultimately, translation of these discoveries into improved treatments, with fundamentally novel mechanisms of action, might require such advances, so that a particular treatment can be matched to a particular genotype or endophenotype. More invasive treatments might also become feasible for severely ill individuals, including deep brain stimulation84 or even viral-mediated gene transfer35,85, to correct abnormalities observed in particular patients. Of course, all of this remains a promissory note. Given past failures to develop non-monoamine-based antidepressants, it is possible that there is something unique about prolonged enhancement of serotonergic or noradrenergic function that causes palliative improvement in a wide range of stress-related disorders including depression and many other conditions. But we reject this nihilistic view on the basis of the extraordinary advances in neurobiology and molecular therapeutics, which make it difficult for us to fully anticipate today improvements that might occur decades from now in psychiatric treatments. We believe that the difficulty of developing non-monoamine-based antidepressants must not obfuscate the importance and eventual feasibility of the goal, given the great clinical need.


References
Manji, H. K., Drevets, W. C. & Charney, D. S. The cellular neurobiology of depression. Nature Med. 7, 541–547 (2001).
Nestler, E. J. et al. Neurobiology of depression. Neuron 34, 13–25 (2002).
Gillespie, C. F. & Nemeroff, C. B. Hypercortisolemia and depression. Psychosom. Med. 67 (Suppl.), S26–S28 (2005).
Charney, D. S. Psychobiological mechanisms of resilience and vulnerability: implications for successful adaptation to extreme stress. Am. J. Psychiatry 161, 195–216 (2004). A comprehensive review of the many neurotransmitters, neuropeptides and hormones, and their receptors, that have a role in an individual's responses to stress. The author argues that our understanding of depression must include a consideration of resilience (resistance to deleterious effects of stress) as well as vulnerability to such effects.
Nestler, E. J. & Carlezon, W. A. Jr. The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry (in the press).
Drevets, W. C. Neuroimaging and neuropathological studies of depression: implications for the cognitive-emotional features of mood disorders. Curr. Opin. Neurobiol. 11, 240–249 (2001).
Bissette, G. et al. Elevated concentrations of CRF in the locus coeruleus of depressed subjects. Neuropsychopharmacology 28, 1328–1335 (2003).
Mayberg, H. S. Positron emission tomography imaging in depression: a neural systems perspective. Neuroimaging Clin. N. Am. 13, 805–815 (2003).
Rajkowska, G. Depression: what we can learn from postmortem studies. Neuroscientist 9, 273–284 (2003).
Morilak, D. A. & Frazer, A. Antidepressants and brain monoaminergic systems: a dimensional approach to understanding their behavioural effects in depression and anxiety disorders. Int. J. Neuropsychopharmacol. 7, 193–218 (2004). Offers a critical evaluation of monoaminergic mechanisms in antidepressant action.
Duman, R. S. Role of neurotrophic factors in the etiology and treatment of mood disorders. Neuromolecular Med. 5, 11–25 (2004). Summarizes the evidence in support of the neurotrophic hypothesis of depression and antidepressant action. In this and other recent reviews, he also discusses several ways to take advantage of this hypothesis for the development of antidepressant agents with novel mechanisms of action, for example, PDE4 inhibitors, and agents directed against neurotrophic signalling proteins.
Sapolsky, R. M. Stress hormones: good and bad. Neurobiol. Dis. 7, 540–542 (2000).
Pariante, C. M. & Miller, A. H. Glucocorticoid receptors in major depression: relevance to pathophysiology and treatment. Biol. Psychiatry 49, 391–404 (2001).
Barden, N. Implication of the hypothalamic–pituitary–adrenal axis in the physiopathology of depression. J. Psychiatry Neurosci. 29, 185–193 (2004).
Korte, S. M., Koohaas, J. M., Wingfield, J. C. & McEwen, B. S. The Darwinian concept of stress: benefits of allostasis and costs of allostatic load and the trade-offs in health and disease. Neurosci. Biobehav. Rev. 29, 3–38 (2005). The authors consider an issue at the heart of stress and depression research today, namely, the necessary and beneficial effects of glucocorticoids, and their potential deleterious effects after periods of prolonged and excessive stress.
de Kloet, E. R., Joels, M. & Holsboer, F. Stress and the brain: from adaptation to disease. Nature Rev. Neurosci. 6, 463–475 (2005). This review offers an excellent discussion of stress physiology, integrating molecular and behavioural data from animal models to provide a clinical perspective.
Dranovsky, A. & Hen, R. Hippocampal neurogenesis: regulation by stress and monoamines. Biol. Psychiatry (in the press).
Walker, K. L., Toufexis, D. J. & Davis, M. Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety. Eur. J. Pharmacol. 163, 199–216 (2003).
Pare, D., Quirk, G. J. & LeDoux, J. E. New vistas on amygdala networks in conditioned fear. J. Neurophysiol. 92, 1–9 (2004).
Keck, M. E., Ohl, F., Holsboer, F. & Muller, M. B. Listening to mutant mice: a spotlight on the role of CRF/CRF receptor systems in affective disorders. Neurosci. Biobehav. Rev. 29, 867–889 (2005). Reviews several lines of mutant mice that have helped to clarify the role of endocrine versus neurotransmitter functions of CRF and the role of CRF 1 and CRF 2 receptors in these responses. For example, selective deletion of non-pituitary CRF 1 receptors in the limbic system is sufficient to suppress the anxiogenic influence of new environments, whereas basal and stress-induced HPA axis activity are unaffected.
Charmandari, E., Tsigos, C. & Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 67, 259–284 (2005).
Bale, T. L. & Vale, W. W. CRF and CRF receptors: role in stress responsivity and other behaviors. Annu. Rev. Pharmacol. Toxicol. 44, 525–557 (2004).
Li, Y. W. et al. The pharmacology of DMP696 and DMP904, non-peptidergic CRF1 receptor antagonists. CNS Drug Rev. 11, 21–52 (2005). Provides a comprehensive synthesis of animal data available with CRF antagonists in depression-related models.
Heinrichs, S. C. & Koob, G. F. Corticotropin-releasing factor in brain: a role in activation, arousal, and affect regulation. J. Pharmacol. Exp. Ther. 311, 427–440 (2004).
Zobel, A. W. et al. Effects of the high-affinity corticotropin-releasing hormone receptor 1 antagonist R121919 in major depression: the first 20 patients treated. J. Psychiatry Res. 34, 171–181 (2000).
Bosker, F. J. et al. Future antidepressants: what is in the pipeline and what is missing? CNS Drugs 18, 705–732 (2004).
Keck, M. E. et al. Reduction of hypothalamic vasopressinergic hyperdrive contributes to clinically relevant behavioral and neuroendocrine effects of chronic paroxetine treatment in a psychopathological rat model. Neuropsychopharmacology 28, 235–243 (2003).
Holmes, A. et al. Neuropeptide systems as novel therapeutic targets for depression and anxiety disorders. Trends Pharmacol. Sci. 24, 580–588 (2003).
Wersinger, S. R. et al. Vasopressin V1b receptor knockout reduces aggressive behavior in male mice. Mol. Psychiatry 7, 975–984 (2002).
Winslow, J. T. & Insel, T. R. Neuroendocrine basis of social recognition. Curr. Opin. Neurobiol. 14, 248–253 (2004).
Boyle, M. P. et al. Acquired deficit of forebrain glucocorticoid receptor produces depression-like changes in adrenal axis regulation and behavior. Proc. Natl Acad. Sci. USA 102, 473–478 (2005). The authors used transgenic mice, in which Cre recombinase is expressed under the control of the calcium/calmodulin-dependent protein kinase II promoter, to generate a forebrain-specific knockdown of the glucocorticoid receptor and show the resulting behavioural and neuroendocrine phenotype.
Wei, Q. et al. Glucocorticoid receptor overexpression in forebrain: a mouse model of increased emotional liability. Proc. Natl Acad. Sci. USA 101, 11851–11856 (2004). Mice with targeted overexpression of glucocorticoid receptors in the forebrain show enhanced negative affective responses in new or stressful situations and increased responsiveness to monoamine-based antidepressants.
Binder, E. B. et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nature Genet. 36, 1319–1325 (2004). This clinical study reports that carriers of a polymorphism in the FKBP5 gene respond faster to antidepressants and have a higher recurrence of depressive episodes than individuals without this polymorphism. FKBP5 encodes a co-chaperone of heat-shock protein 90; the variant causes the glucocorticoid receptor to exhibit a higher affinity for cortisol.
Kaufer, D. et al. Restructuring the neuronal stress response with anti-glucocorticoid gene delivery. Nature Neurosci. 7, 947–953 (2004). Shows, by use of viral-mediated gene transfer in rat, that genetic modification of glucocorticoid receptors diminishes the deleterious effects of glucocorticoids both in vitro and in vivo.
Akama, K. T. & McEwen, B. S. Gene therapy to bet on: protecting neurons from stress hormones. Trends Pharmacol. Sci. 26, 169–172 (2005).
Flores, H. F. et al. Clinical and biological effects of mifepristone treatment for psychotic depression. Neuropharmacology 14 Sep 2005 (10.1038/sj.npp.1300884). The first report of a double-blind placebo-controlled study showing the efficacy of mifepristone in the treatment of psychotic depression. Alterations in the HPA axis were also observed.
Adell, A. et al. Strategies for producing faster acting antidepressants. Drug Discov. Today 10, 578–585 (2005).
Jahn, H. et al. Metyrapone as additive treatment in major depression: a double-blind and placebo-controlled trial. Arch. Gen. Psychiatry 61, 1235–1244 (2004).
Adell, A. Antidepressant properties of substance P antagonists: relationship to monoaminergic mechanisms? Curr. Drug Targets CNS Neurol. Disord. 3, 113–121 (2004).
Rupniak, N. M. J. et al. Comparison of the phenotype of NK1R−/− mice with pharmacological blockade of the substance P (NK1) receptor in assays for antidepressant and anxiolytic drugs. Behav. Pharmacol. 12, 497–508 (2001).
Ebner, K., Rupniak, N. M., Saria, A. & Singewald, N. Substance P in the medial amygdala: emotional stress-sensitive release and modulation of anxiety-related behavior in rats. Proc. Natl Acad. Sci. USA 101, 4280–4285 (2004).
Kramer, M. S. et al. Distinct mechanism for antidepressant activity by blockade of central substance P receptors. Science 281, 1640–1645 (1998).
Blier, P. et al. Impact of substance P receptor antagonism on the serotonin and norepinephrine systems: relevance to the antidepressant/anxiolytic response. J. Psychiatry Neurosci. 29, 208–218 (2004). Summarizes the potential interactions between the substance P–NK 1 system and monoaminergic systems in the brain, based on genetic and pharmacological models, as they relate to antidepressant and anxiolytic drug mechanisms.
Ryckmans, T. et al. First dual NK1 antagonists-serotonin reuptake inhibitors: synthesis and SAR of a new class of potential antidepressants. Bioorg. Med. Chem. Lett. 12, 261–264 (2002).
Morcuende, S. et al. Increased neurogenesis and brain-derived neurotrophic factor in neurokinin-1 receptor gene knockout mice. Eur. J. Neurosci. 18, 1828–1836 (2003).
Guest, P. C. et al. Mechanisms of action of the antidepressants fluoxetine and the substance P antagonist L-000760735 are associated with altered neurofilaments and synaptic remodeling. Brain Res. 1002, 1–10 (2004).
Guiard, B. P. et al. Blockade of substance P (neurokinin 1) receptors enhances extracellular serotonin when combined with a selective serotonin reuptake inhibitor: an in vivo microdialysis study in mice. J. Neurochem. 89, 54–63 (2004).
Musazzi, L., Perez, J., Hunt, S. P., Racagni, G. & Popoli, M. Changes in signaling pathways regulating neuroplasticity induced by neurokinin 1 receptor knockout. Eur. J. Neurosci. 21, 1370–1378 (2005).
van der Hart, M. G. et al. Chronic psychosocial stress in tree shrews: effect of the substance P (NK1 receptor) antagonist L-760735 and clomipramine on endocrine and behavioral parameters. Psychopharmacology (Berl.) (in the press). Compares the effects of substance P antagonists and tricyclic antidepressants on neuroendocrine and behavioural parameters in a unique primate model of chronic psychosocial stress.
Duman, R. S., Heninger, G. R. & Nestler, E. J. A molecular and cellular hypothesis of depression. Arch. Gen. Psychiatry 54, 597–606 (1997).
Chen, B. et al. Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol. Psychiatry 50, 260–265 (2001).
Shirayama, Y. et al. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J. Neurosci. 22, 3251–3261 (2002).
Monteggia, L. M. et al. Essential role of BDNF in adult hippocampal function. Proc. Natl Acad. Sci. USA 101, 10827–10832 (2004). Used an inducible, cell-targeted knockout system to obtain one of the best demonstrations so far that forebrain BDNF is necessary for antidepressant efficacy in the forced swim test.
Eisch, A. J. BDNF in the ventral midbrain–nucleus accumbens pathway: a role in depression. Biol. Psychiatry 54, 994–1005 (2003).
Berton, O. et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science (in the press). Chronic social defeat stress causes a long-lasting abnormality in social approach-avoidance behaviour, which is reversed by chronic (but not acute) antidepressant administration. Selective deletion of BDNF in the ventral tegmental area prevents this behavioural abnormality of defeat stress, and induces changes in gene expression in the nucleus accumbens similar to those seen with chronic antidepressant use.
Turner, C. A., Akil, A., Watson, S. J. & Evans, S. J. The fibroblast growth factor system and mood disorders. Biol. Psychiatry (in the press). Summarizes recent evidence, much of it from DNA microarray analysis of human brain tissue taken at postmortem, for abnormalities in the FGF system in depression.
Simen, B. B., Duman, C. H., Simen, A. A. & Duman, R. S. TNFα signaling in depression and anxiety: behavioral consequences of individual receptor targeting. Biol. Psychiatry (in the press). One of the first studies to characterize depression-like behaviour in animals lacking TNFα. These types of analysis of the large number of mutants available in the cytokine field will help us to clarify the influence of various cytokines and their receptors on depression-like behaviours and antidepressant responses.
Chen, A. C., Shirayama, Y., Shin, K. H., Neve, R. L. & Duman, R. S. Expression of the cAMP response element binding protein (CREB) in hippocampus produces an antidepressant effect. Biol. Psychiatry 49, 753–762 (2001).
Tully, T., Bourtchouladze, R., Scott, R. & Tallman, J. Targeting the CREB pathway for memory enhancers. Nature Rev. Drug Discov. 2, 267–277 (2003).
Mayford, M. & Kandel, E. R. Genetic approaches to memory storage. Trends Genet. 15, 463–470 (1999).
Carlezon, W. A. Jr, Duman, R. S. & Nestler, E. J. The many faces of CREB. Trends Pharmacol. Sci. 28, 436–445 (2005).
Ramos, B. P. et al. Dysregulation of protein kinase A signaling in the aged prefrontal cortex: new strategy for treating age-related cognitive decline. Neuron 40, 835–845 (2003).
Sanacora, G., Rothman, D. L., Mason, G. & Krystal, J. H. Clinical studies implementing glutamate neurotransmission in mood disorders. Ann. NY Acad. Sci. 1003, 292–308 (2003).
Paul, I. A. & Skolnick, P. Glutamate and depression: clinical and preclinical studies. Ann. NY Acad. Sci. 1003, 250–272 (2003).
Miyamoto, Y. et al. Lower sensitivity to stress and altered monoaminergic neuronal function in mice lacking the NMDA receptor ε4 subunit. J. Neurosci. 22, 2335–2342 (2002).
Alt, A., Witkin, J. M. & Bleakman, D. AMPA receptor potentiators as novel antidepressants. Curr. Pharm. Des. 11, 1511–1527 (2005). Discusses the use of positive allosteric modulators of AMPA glutamate receptors as putative antidepressant agents.
Swanson, C. J. et al. Metabotropic glutamate receptors as novel targets for anxiety and stress disorders. Nature Rev. Drug Discov. 4, 131–144 (2005).
Saito, Y. et al. Molecular characterization of the melanin-concentrating-hormone receptor. Nature 400, 265–269 (1999).
Georgescu, D. et al. The neuropeptide MCH controls feeding behavior via a novel hypothalamic-limbic circuit. J. Neurosci. 25, 2933–2940 (2005). Using molecular and pharmacological tools, this provides direct evidence that MCH, acting in the nucleus accumbens, exerts a prodepression-like effect, with antidepressant-like effects exerted by MCH receptor antagonism in this brain region.
Borowsky, B. et al. Antidepressant, anxiolytic and anorectic effects of a melanin-concentrating hormone-1 receptor antagonist. Nature Med. 8, 825–830 (2002).
Roy, M. et al. Genetic inactivation of melanin-concentrating hormone receptor subtype 1 (MCHR1) in mice exerts anxiolytic-like behavioral effects. Neuropsychopharmcology (in the press). One of several recent reports of the phenotype of MCH receptor-knockout mice. The other articles consider metabolic abnormalities of these knockouts, whereas this study shows a reduction in anxiety-like behaviours. Depression-like behaviour has not yet been reported in MCH receptor-knockouts.
Winsky-Sommerer, R. et al. Interaction between the corticotropin-releasing factor system and hypocretins (orexins): a novel circuit mediating stress response. J. Neurosci. 24, 11439–11448 (2004).
Takahashi, J. S. Finding new clock components: past and future. J. Biol. Rhythms 19, 339–347 (2004).
Reppert, S. M. & Weaver, D. R. Coordination of circadian timing in mammals. Nature 418, 935–941 (2002).
McClung, C. et al. Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc. Natl Acad. Sci. USA 102, 9377–9381 (2005). Demonstrates that mice lacking functional clock protein show enhanced behavioural responses to cocaine and enhanced firing of ventral tegmental area dopamine neurons. This is one of an increasing number of reports that circadian genes act outside the SCN to regulate complex behaviour.
Uz, T. et al. Effect of fluoxetine and cocaine on the expression of clock genes in the mouse hippocampus and striatum. Neuroscience (in the press).
McClung, C. A. et al. Disruption of the clock gene in mice induces a manic-like state. Soc. Neurosci. Abstr. 793.20 (2005).
Garcia, J. A. et al. Impaired cued and contextual memory in NPAS2-deficient mice. Science 288, 2226–2230 (2000).
Dudley, C. A. et al. Altered patterns of sleep and behavioral adaptability in NPAS2-deficient mice. Science 301, 379–383 (2003). This and related studies by the same authors document a role for NPAS2, a clock-like transcription factor, in circadian variations in feeding, sleeping and other complex behaviours.These effects of NPAS2 are independent of the SCN and apparently mediated via forebrain regions.
Manji, H. K. et al. Enhancing neuronal plasticity and cellular resilience to develop novel, improved therapeutics for difficult-to-treat depression. Biol. Psychiatry 53, 707–742 (2003).
Li, X., Bijur, G. N. & Jope, R. S. Glycogen synthase kinase-3β, mood stabilizers, and neuroprotection. Bipolar Disord. 4, 137–144 (2002).
Markou, A. (ed.) Animal models of depression and antidepressant activity. Neurosci. Biobehav. Rev. 29, 501–909 (2005). A recent issue of Neuroscience and Biobehavioral Reviews that is devoted entirely to animal models of depression. It includes numerous outstanding and up-to-date reviews of diverse behavioural paradigms used to study depression-like behaviour and antidepressant action in animals.
Nestler, E. J. et al. Preclinical models: status of basic research in depression. Biol. Psychiatry 52, 503–528 (2002).
Mayberg, H. S. et al. Deep brain stimulation for treatment-resistant depression. Neuron 45, 651–660 (2005). This study of six patients with severe depression showed that most experienced some benefit on deep brain stimulation of a region of cingulate cortex, which showed abnormal functioning in brain imaging investigations of these patients.
Sapolsky, R. Gene therapy for psychiatric disorders. Am. J. Psychiatry 160, 208–220 (2003).
Diagnostic Statistical Manual of Mental Disorders (American Psychiatric Press, Washington, DC, 2000).
Mague, S. D. et al. Antidepressant-like effects of κ-opioid receptor antagonists in the forced swim test in rats. J. Pharmacol. Exp. Ther. 305, 323–330 (2003).
McLaughlin, J. P., Marton-Popovici, M. & Chavkin, C. κ opioid receptor antagonism and prodynorphin gene disruption block stress-induced behavioral responses. J. Neurosci. 23, 5674–5683 (2003). References 87 and 88 were two of the first to demonstrate antidepressant-like effects of κ opioid antagonists after systemic administration.
Shippenberg, T. S. & Rea, W. Sensitization to the behavioral effects of cocaine: modulation by dynorphin and κ-opioid receptor agonists. Pharmacol. Biochem. Behav. 57, 449–455 (1997).
Viveros, M. P., Marco, E. M. & File, S. E. Endocannabinoid system and stress and anxiety responses. Pharmacol. Biochem. Behav. 81, 331–342 (2005).
Haller, J. et al. CB1 cannabinoid receptors mediate anxiolytic effects: convergent genetic and pharmacological evidence with CB1-specific agents. Behav. Pharmacol. 15, 299–304 (2004).
Kathuria, S. et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nature Med. 9, 76–81 (2003). Shows that genetic ablation of one of the enzymes that degrade the endogenous cannabinoid ligand anandamide exerts anxiolytic-like effects in rodents.
Griebel, G., Stemmelin, J. & Scatton, B. Effects of the cannabinoid CB1 receptor antagonist rimonabant in models of emotional reactivity in rodents. Biol. Psychiatry 57, 261–267 (2005). Characterizes the effects of the CB 1 antagonist rimonabant, now in clinical trials for obesity, on emotional behaviour relevant to anxiety- and depression-like symptoms.
Gobbi, G. et al. Antidepressant-like activity and modulation of brain monoaminergic transmission by blockade of anandamide hydrolysis. Proc. Natl Acad. Sci. USA 13 Dec 2005 (10.1073/pnas.0509591102) The authors show antidepressant-like effects of an inhibitor of one of the enzymes that degrade anandamide, a major endogenous ligand of the CB 1 receptor.
Dunn, A. J., Swiergiel, A. H. & de Beaurepaire, R. Cytokines as mediators of depression: What can we learn from animal studies? Neurosci. Biobehav. Rev. 29, 891–909 (2005). Provides an excellent review of the actions of cytokines in the brain and their possible relevance to depression and antidepressant treatments.
Castanon, N., Medina, C., Mormede, C. & Dantzer, R. Chronic administration of tianeptine balances lipopolysaccharide-induced expression of cytokines in the spleen and hypothalamus of rats. Psychoneuroendocrinology 29, 778–790 (2004).
Strle, K. et al. Interleukin-10 in the brain. Crit. Rev. Immunol. 21, 427–449 (2001).
Barden, N. et al. Antidepressant action of agomelatine (S 20098) in a transgenic mouse model. Prog. Neuropsychopharmacol. Biol. Psychiatry 29, 908–916 (2005).
Holmes, A. et al. Phenotypic assessment of galanin overexpressing and galanin receptor R1 knockout mice in the tail suspension test for depression-related behavior. Psychopharmacology 178, 276–285 (2005).
Levenson, J. M. & Sweatt, J. D. Epigenetic mechanisms in memory formation. Nature Rev. Neurosci. 6, 108–118 (2005). An excellent review of an evolving literature that demonstrates the influence of chromatin remodelling on synaptic plasticity and learning and memory.
Alarcon, J. M. et al. Chromatin acetylation, memory, and LTP are impaired in CBP+/− mice: a model for the cognitive deficit in Rubinstein–Taybi syndrome and its amelioration. Neuron 42, 947–959 (2004).
Tsankova, N. M., Kumar, A. & Nestler, E. J. Histone modifications at gene promoter regions in rat hippocampus after acute and chronic electroconvulsive seizures. J. Neurosci. 24, 5603–5610 (2004).
Kumar, A. et al. Chromatin remodeling is a key mechanism underlying cocaine-induced plasticity in striatum. Neuron 48, 303–314 (2005).
Tsankova, N. M., Berton, O. & Nestler, E. J. Stable changes in histone modifications at the BDNF gene in hippocampus in depression and antidepressant action: possible role for HDAC5. Soc. Neurosci. Abstr. 638.13 (2005).
Weaver, I. C. et al. Epigenetic programming by maternal behavior. Nature Neurosci. 7, 847–854 (2004). The authors demonstrate that early maternal separation causes hypermethylation of the glucocorticoid receptor gene, which underlies several behavioural and neuroendocrine abnormalities seen in the animals that survive to adulthood.
Tomasz Matys, T. et al. Tissue plasminogen activator promotes the effects of corticotropin-releasing factor on the amygdala and anxiety-like behavior. Proc. Natl Acad. Sci. USA 101, 16345–16350 (2004).
Acknowledgements
Preparation of this review was supported by grants from the National Institute of Mental Health, USA.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
E.J.N. receives consultancy income from several pharmaceutical and biotechnology companies with active antidepressant drug discovery programmes. He is Chair of a neuroscience Scientific Advisory Board of Eli Lilly, USA, and a member of the Scientific Advisory Boards of Neurogen, Helicon, Neurologix, Neuromolecular, RxGen and Intracellular Therapies (all USA). He is also co-founder and Scientific Advisory Board Chair of PsychoGenics, USA.
Related links
Glossary
- Anhedonia
-
Decreased interest in, and ability to experience, pleasure. A common symptom of depression.
- Monoamine neurotransmitters
-
Small molecule neurotransmitters that contain a single amine group. Monoamines include dopamine, serotonin, noradrenaline and adrenaline, and histamine is sometimes included in this group of neurotransmitters as well.
- Electroconvulsive therapy
-
(ECT). Repeated generalized seizures, induced electrically, as a treatment for depression. A form of somatic therapy.
- Somatic therapies
-
Refers to non-medication, non-psychotherapy treatments for depression. Such therapies include ECT and several more experimental treatments such as magnetic stimulation (transcranial magnetic stimulation, TMS, and magnetic seizures) and vagal nerve stimulation.
- Tricyclic antidepressants
-
Refers to a group of structurally related compounds that were developed in the 1950s and later shown to possess antidepressant activity in humans. Prototypical tricyclic antidepressants include amitriptyline, imipramine and desipramine.
- Atypical antidepressants
-
Clinically effective antidepressants for which the mechanisms of action are unknown (for example, bupropion, mirtazapine and tianeptine).
- Stress models
-
The application of any of several aversive or nociceptive stimuli (physical or psychological stressors) to an animal. Examples include restraint stress, foot shock, social defeat and chronic mild stress.
- Monoaminergic depletion
-
Tryptophan depletion lowers serotonin levels in the brain. This treatment increases anxiety-related behaviours in rodents and increases immobility in the forced swim test. No clear effects of antidepressants have been reported.
- Olfactory bulbectomy
-
Causes degeneration of neurons connecting the olfactory bulbs to the limbic system. This is associated with several behavioural abnormalities that can be normalized by chronic, but not acute, antidepressant treatment.
- Hyponeophagia
-
(Novelty-suppressed feeding). Inhibition of feeding produced by exposure to a novel environment provides an anxiety-related measure that is sensitive to the effects of chronic, but not acute, antidepressant treatment. The test also responds to anxiolytic drugs, such as benzodiazepines.
- Learned helplessness
-
Reduced escape behaviour in response to stress after prior exposures to unavoidable stressors. Responds to acute or subchronic antidepressant administration.
- Chronic mild or unpredictable stress
-
Involves relatively prolonged exposure to various relatively mild stressors. Reported to induce an anhedonia-like state, which can be reversed by chronic antidepressant treatment. However, these findings have not been replicated by all laboratories.
- Chaperone proteins
-
Proteins that bind to other proteins to regulate their folding, trafficking among intracellular compartments, and degradation.
- Cushing syndrome
-
Medical consequences of hypersecretion of glucocorticoids from the adrenal cortex, which can be caused by several conditions.
- Distress vocalizations
-
Ultrasonic vocalizations produced by young rodents lost outside the nest or by adults in aversive contexts. Acute treatment with several types of antidepressant inhibits the production of these calls.
- Forced swim test
-
Rodents immersed in a vessel of water develop an immobile posture after initial struggling. Most antidepressants, administered acutely before the test, reverse the immobility and promote struggling. Advantages of this technique include low cost, high throughput and predictive validity; disadvantages include the fact that acute antidepressant administration, which is not effective in human depression, is effective in the test
- Social defeat
-
Prolonged exposure to an aggressor causes several depression-like behavioural abnormalities in animals, which can be reversed by chronic, but not acute, antidepressant treatment. Social defeat is an example of chronic psychosocial stress.
- Psychotomimetic drug
-
A drug that induces psychosis. Prototypical examples include NMDA receptor antagonists (for example, phencyclidine and ketamine) and psychostimulants administered repeatedly at high doses (for example, amphetamine).
- Tail suspension test
-
Mice suspended by their tails develop an immobile posture after initial struggling. Acute administration of most antidepressants before the test reverses immobility and promotes struggling. Advantages of this technique include low cost, high throughput and predictive validity; disadvantages include the fact that acute antidepressant administration, which is not effective in human depression, is effective in the test.
- Brain stimulation reward
-
Rodents will work (press a lever) to pass electric current into specific brain areas. A change in the threshold current for such intracranial self-stimulation is reported to provide a measure of affective state, with an increase in threshold current reflecting a depressed affect.
Rights and permissions
About this article
Cite this article
Berton, O., Nestler, E. New approaches to antidepressant drug discovery: beyond monoamines. Nat Rev Neurosci 7, 137–151 (2006). https://doi.org/10.1038/nrn1846
Issue Date:
DOI: https://doi.org/10.1038/nrn1846
This article is cited by
-
Akkermansia muciniphila Improves Depressive-Like Symptoms by Modulating the Level of 5-HT Neurotransmitters in the Gut and Brain of Mice
Molecular Neurobiology (2024)
-
Egg consumption reduces the risk of depressive symptoms in the elderly: findings from a 6-year cohort study
BMC Psychiatry (2023)
-
Impact of St. John’s wort extract Ze 117 on stress induced changes in the lipidome of PBMC
Molecular Medicine (2023)
-
A computational analysis of mouse behavior in the sucrose preference test
Nature Communications (2023)
-
Dextromethorphan-Bupropion for the Treatment of Depression: A Systematic Review of Efficacy and Safety in Clinical Trials
CNS Drugs (2023)
