
Key Points
-
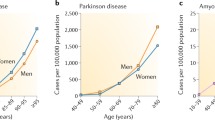
Aging is the single major risk factor for the development of human neurodegenerative diseases. Typically, familial mutation-linked neurodegeneration emerges during the fifth decade of life, whereas the onset of sporadic neurodegenerative disease is usually during the seventh decade or later.
-
Human neurodegenerations share a tight mechanistic link to toxic protein aggregation.
-
Insulin/insulin-like growth factor 1 (IGF1) signalling (IIS) is a prominent lifespan and stress-resistance regulator.
-
Altering aging by reducing IIS protects model animals from the toxicity that is associated with the aggregation of disease-linked proteins.
-
Two opposing mechanisms mediate the counter-toxic effect of a reduction in IIS: disaggregation and active aggregation. These protective activities are regulated by heat-shock factor 1 (HSF1) and by the IIS downstream transcription factor DAF-16, respectively.
-
Contradictory data indicate that IGF1 infusion protects rodents from the aggregation that is associated with the Alzheimer's-disease-linked peptide amyloid-β.
-
A model that predicts an optimal IIS level could accommodate this apparent paradox.
Abstract
Distinct human neurodegenerative diseases share remarkably similar temporal emergence patterns, even though different toxic proteins are involved in their onset. Typically, familial neurodegenerative diseases emerge during the fifth decade of life, whereas sporadic cases do not exhibit symptoms earlier than the seventh decade. Recently, mechanistic links between the aging process and toxic protein aggregation, a common hallmark of neurodegenerative diseases, have been revealed. The insulin/insulin-like growth factor 1 (IGF1) signalling pathway — a lifespan, metabolism and stress-resistance regulator — links neurodegeneration to the aging process. Thus, although a reduction of insulin signalling can result in diabetes, its reduction can also increase longevity and delay the onset of protein-aggregation-mediated toxicity. Here we review this apparent paradox and delineate the therapeutic potential of manipulating the insulin/IGF1 signalling pathway for the treatment of neurodegenerative diseases.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

References
Reichel, W. The biology of aging. J. Am. Geriatr. Soc. 14, 431–436 (1966).
Bishop, N. A. & Guarente, L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nature Rev. Genet. 8, 835–844 (2007).
Mair, W. & Dillin, A. Aging and survival: the genetics of life span extension by dietary restriction. Annu. Rev. Biochem. 77, 727–754 (2008).
Kenyon, C., Chang, J., Gensch, E., Rudner, A. & Tabtiang, R. A C. elegans mutant that lives twice as long as wild type. Nature 366, 461–464 (1993).
Giannakou, M. E. & Partridge, L. Role of insulin-like signalling in Drosophila lifespan. Trends Biochem. Sci. 32, 180–188 (2007).
Tatar, M. et al. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 292, 107–110 (2001).
Holzenberger, M. et al. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature 421, 182–187 (2003). This study established that the longevity and stress-resistance effects of reduced IIS are conserved from invertebrates to mammals.
Russell, S. J. & Kahn, C. R. Endocrine regulation of ageing. Nature Rev. Mol. Cell Biol. 8, 681–691 (2007).
Taguchi, A., Wartschow, L. M. & White, M. F. Brain IRS2 signaling coordinates life span and nutrient homeostasis. Science 317, 369–372 (2007).
Dillin, A. et al. Rates of behavior and aging specified by mitochondrial function during development. Science 298, 2398–2401 (2002).
Lee, S. S. et al. A systematic RNAi screen identifies a critical role for mitochondria in C. elegans longevity. Nature Genet. 33, 40–48 (2003).
Taguchi, A. & White, M. F. Insulin-like signaling, nutrient homeostasis, and life span. Annu. Rev. Physiol. 70, 191–212 (2008).
Balch, W. E., Morimoto, R. I., Dillin, A. & Kelly, J. W. Adapting proteostasis for disease intervention. Science 319, 916–919 (2008).
Aguilaniu, H., Gustafsson, L., Rigoulet, M. & Nystrom, T. Asymmetric inheritance of oxidatively damaged proteins during cytokinesis. Science 299, 1751–1753 (2003).
Hebert, D. N. & Molinari, M. In and out of the ER: protein folding, quality control, degradation, and related human diseases. Physiol. Rev. 87, 1377–1408 (2007).
Gidalevitz, T., Ben-Zvi, A., Ho, K. H., Brignull, H. R. & Morimoto, R. I. Progressive disruption of cellular protein folding in models of polyglutamine diseases. Science 311, 1471–1474 (2006). In this article the authors reported that the expression of one aggregation-prone polyQ protein can destabilize the genome and lead to the aggregation of other proteins.
Kopito, R. R. & Ron, D. Conformational disease. Nature Cell Biol. 2, E207–E209 (2000).
Selkoe, D. J. Folding proteins in fatal ways. Nature 426, 900–904 (2003).
Bates, G. Huntingtin aggregation and toxicity in Huntington's disease. Lancet 361, 1642–1644 (2003).
Caughey, B. & Lansbury, P. T. Protofibrils, pores, fibrils, and neurodegeneration: separating the responsible protein aggregates from the innocent bystanders. Annu. Rev. Neurosci. 26, 267–298 (2003).
Haass, C. & Selkoe, D. J. Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer's amyloid b-peptide. Nature Rev. Mol. Cell Biol. 8, 101–112 (2007).
Forloni, G. Neurotoxicity of beta-amyloid and prion peptides. Curr. Opin. Neurol. 9, 492–500 (1996).
Kim, S. J. & Linden, D. J. Ubiquitous plasticity and memory storage. Neuron 56, 582–592 (2007).
Shankar, G. M. et al. Amyloid-β protein dimers isolated directly from Alzheimer's brains impair synaptic plasticity and memory. Nature Med. 22 Jun 2008 (doi:10.1038/nm1782).
Townsend, M., Shankar, G. M., Mehta, T., Walsh, D. M. & Selkoe, D. J. Effects of secreted oligomers of amyloid β-protein on hippocampal synaptic plasticity: a potent role for trimers. J. Physiol. 572, 477–492 (2006).
Arrasate, M., Mitra, S., Schweitzer, E. S., Segal, M. R. & Finkbeiner, S. Inclusion body formation reduces levels of mutant huntingtin and the risk of neuronal death. Nature 431, 805–810 (2004).
Behrends, C. et al. Chaperonin TRiC promotes the assembly of polyQ expansion proteins into nontoxic oligomers. Mol. Cell 23, 887–897 (2006). Along with reference 28, this work indicated that chaperones, which are known to disrupt protein aggregates, switch to actively create aggregates of high molecular mass when the concentration of the aggregating protein is high. These papers thus established the idea that active aggregation can be protective.
Shorter, J. & Lindquist, S. Hsp104 catalyzes formation and elimination of self-replicating Sup35 prion conformers. Science 304, 1793–1797 (2004).
Amaducci, L. & Tesco, G. Aging as a major risk for degenerative diseases of the central nervous system. Curr. Opin. Neurol. 7, 283–286 (1994).
Brignull, H. R., Morley, J. F. & Morimoto, R. I. The stress of misfolded proteins: C. elegans models for neurodegenerative disease and aging. Adv. Exp. Med. Biol. 594, 167–189 (2007).
Jankowsky, J. L. et al. Transgenic mouse models of neurodegenerative disease: opportunities for therapeutic development. Curr. Neurol. Neurosci. Rep. 2, 457–464 (2002).
Cohen, E., Bieschke, J., Perciavalle, R. M., Kelly, J. W. & Dillin, A. Opposing activities protect against age-onset proteotoxicity. Science 313, 1604–1610 (2006).
Hsu, A. L., Murphy, C. T. & Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 300, 1142–1145 (2003). This paper showed that HSF-1 is crucial for enabling reduced IIS to extend lifespan.
Morley, J. F., Brignull, H. R., Weyers, J. J. & Morimoto, R. I. The threshold for polyglutamine-expansion protein aggregation and cellular toxicity is dynamic and influenced by aging in Caenorhabditis elegans. Proc. Natl Acad. Sci. USA 99, 10417–10422 (2002). This seminal study indicated that the minimal number of polyQ repeats that is needed to enable aggregation declines with age. It also showed that a reduction in IIS protects worms from polyQ aggregation.
Kimura, K. D., Tissenbaum, H. A., Liu, Y. & Ruvkun, G. daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277, 942–946 (1997).
Morris, J. Z., Tissenbaum, H. A. & Ruvkun, G. A phosphatidylinositol-3-OH kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 382, 536–539 (1996).
Kops, G. J. et al. Direct control of the Forkhead transcription factor AFX by protein kinase B. Nature 398, 630–634 (1999).
Paradis, S. & Ruvkun, G. Caenorhabditis elegans Akt/PKB transduces insulin receptor-like signals from AGE-1 PI3 kinase to the DAF-16 transcription factor. Genes Dev. 12, 2488–2498 (1998).
Ogg, S. & Ruvkun, G. The C. elegans PTEN homolog, DAF-18, acts in the insulin receptor-like metabolic signaling pathway. Mol. Cell 2, 887–893 (1998).
Henderson, S. T. & Johnson, T. E. daf-16 integrates developmental and environmental inputs to mediate aging in the nematode Caenorhabditis elegans. Curr. Biol. 11, 1975–1980 (2001).
Lee, R. Y., Hench, J. & Ruvkun, G. Regulation of C. elegans DAF-16 and its human ortholog FKHRL1 by the daf-2 insulin-like signaling pathway. Curr. Biol. 11, 1950–1957 (2001).
Murphy, C. T. et al. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424, 277–283 (2003).
Kenyon, C. The plasticity of aging: insights from long-lived mutants. Cell 120, 449–460 (2005).
Bluher, M., Kahn, B. B. & Kahn, C. R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 299, 572–574 (2003).
Suh, Y. et al. Functionally significant insulin-like growth factor I receptor mutations in centenarians. Proc. Natl Acad. Sci. USA 105, 3438–3442 (2008). This report indicated that reduced IGF signalling is typical in the offspring of centenarians and suggested that the regulation of lifespan by IGF signalling is conserved in humans. It also showed that high IGF plasma levels can lead to reduced IGF signalling, supporting the idea that feedback loops regulate the IIS pathway.
Morley, J. F. & Morimoto, R. I. Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol. Biol. Cell 15, 657–664 (2004).
Liu, X. D., Liu, P. C., Santoro, N. & Thiele, D. J. Conservation of a stress response: human heat shock transcription factors functionally substitute for yeast HSF. EMBO J. 16, 6466–6477 (1997).
Rabindran, S. K., Haroun, R. I., Clos, J., Wisniewski, J. & Wu, C. Regulation of heat shock factor trimer formation: role of a conserved leucine zipper. Science 259, 230–234 (1993).
Walker, G. A., Thompson, F. J., Brawley, A., Scanlon, T. & Devaney, E. Heat shock factor functions at the convergence of the stress response and developmental pathways in Caenorhabditis elegans. FASEB J. 17, 1960–1962 (2003).
Sarge, K. D., Murphy, S. P. & Morimoto, R. I. Activation of heat shock gene transcription by heat shock factor 1 involves oligomerization, acquisition of DNA-binding activity, and nuclear localization and can occur in the absence of stress. Mol. Cell. Biol. 13, 1392–1407 (1993).
Baird, N. A., Turnbull, D. W. & Johnson, E. A. Induction of the heat shock pathway during hypoxia requires regulation of heat shock factor by hypoxia-inducible factor-1. J. Biol. Chem. 281, 38675–38681 (2006).
Reinke, H. et al. Differential display of DNA-binding proteins reveals heat-shock factor 1 as a circadian transcription factor. Genes Dev. 22, 331–345 (2008).
Singh, V. & Aballay, A. Heat-shock transcription factor (HSF)-1 pathway required for Caenorhabditis elegans immunity. Proc. Natl Acad. Sci. USA 103, 13092–13097 (2006).
Bishop, N. A. & Guarente, L. Two neurons mediate diet-restriction-induced longevity in C. elegans. Nature 447, 545–549 (2007).
Tullet, J. M. et al. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. elegans. Cell 132, 1025–1038 (2008).
McElwee, J., Bubb, K. & Thomas, J. H. Transcriptional outputs of the Caenorhabditis elegans forkhead protein DAF-16. Aging Cell 2, 111–121 (2003).
Halaschek-Wiener, J. et al. Analysis of long-lived C. elegans daf-2 mutants using serial analysis of gene expression. Genome Res. 15, 603–615 (2005).
Oh, S. W. et al. Identification of direct DAF-16 targets controlling longevity, metabolism and diapause by chromatin immunoprecipitation. Nature Genet. 38, 251–257 (2006).
Dong, M. Q. et al. Quantitative mass spectrometry identifies insulin signaling targets in C. elegans. Science 317, 660–663 (2007).
Tonkiss, J. & Calderwood, S. K. Regulation of heat shock gene transcription in neuronal cells. Int. J. Hyperthermia 21, 433–444 (2005).
Hartl, F. U. & Hayer-Hartl, M. Molecular chaperones in the cytosol: from nascent chain to folded protein. Science 295, 1852–1858 (2002).
Barral, J. M., Broadley, S. A., Schaffar, G. & Hartl, F. U. Roles of molecular chaperones in protein misfolding diseases. Semin. Cell Dev. Biol. 15, 17–29 (2004).
Howitz, K. T. et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 425, 191–196 (2003).
Wood, J. G. et al. Sirtuin activators mimic caloric restriction and delay ageing in metazoans. Nature 430, 686–689 (2004).
Valenzano, D. R. et al. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short-lived vertebrate. Curr. Biol. 16, 296–300 (2006).
Parker, J. A. et al. Resveratrol rescues mutant polyglutamine cytotoxicity in nematode and mammalian neurons. Nature Genet. 37, 349–350 (2005). This study showed that altering aging with the drug Resveratrol protects PolyQ-expressing neurons from protein aggregation. It thus linked the aging process and protein-aggregation neurotoxicity.
Tissenbaum, H. A. & Guarente, L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature 410, 227–230 (2001).
Link, C. Expression of human β-amyloid peptide in transgenic Caenorhabditis elegans. Proc. Natl Acad. Sci. USA 92, 9368–9372 (1995).
Humbert, S. et al. The IGF-1/Akt pathway is neuroprotective in Huntington's disease and involves Huntingtin phosphorylation by Akt. Dev. Cell 2, 831–837 (2002).
Yamamoto, A., Cremona, M. L. & Rothman, J. E. Autophagy-mediated clearance of huntingtin aggregates triggered by the insulin-signaling pathway. J. Cell Biol. 172, 719–731 (2006).
Carro, E., Trejo, J. L., Gomez-Isla, T., LeRoith, D. & Torres-Aleman, I. Serum insulin-like growth factor I regulates brain amyloid-β levels. Nature Med. 8, 1390–1397 (2002).
Carro, E. et al. Therapeutic actions of insulin-like growth factor I on APP/PS2 mice with severe brain amyloidosis. Neurobiol. Aging 27, 1250–1257 (2006).
Gasparini, L. & Xu, H. Potential roles of insulin and IGF-1 in Alzheimer's disease. Trends Neurosci. 26, 404–406 (2003).
Bonkowski, M. S., Rocha, J. S., Masternak, M. M., Al Regaiey, K. A. & Bartke, A. Targeted disruption of growth hormone receptor interferes with the beneficial actions of calorie restriction. Proc. Natl Acad. Sci. USA 103, 7901–7905 (2006).
Tomlinson, D. R. & Gardiner, N. J. Glucose neurotoxicity. Nature Rev. Neurosci. 9, 36–45 (2008).
Zick, Y. Ser/Thr phosphorylation of IRS proteins: a molecular basis for insulin resistance. Sci. STKE pe4 (2005).
George, S. et al. A family with severe insulin resistance and diabetes due to a mutation in AKT2. Science 304, 1325–1328 (2004).
Biddinger, S. B. et al. Hepatic insulin resistance is sufficient to produce dyslipidemia and susceptibility to atherosclerosis. Cell. Metab. 7, 125–134 (2008).
Brown, M. S. & Goldstein, J. L. Selective versus total insulin resistance: a pathogenic paradox. Cell. Metab. 7, 95–96 (2008).
Michael, M. D. et al. Loss of insulin signaling in hepatocytes leads to severe insulin resistance and progressive hepatic dysfunction. Mol. Cell 6, 87–97 (2000).
Craft, S. et al. Cerebrospinal fluid and plasma insulin levels in Alzheimer's disease: relationship to severity of dementia and apolipoprotein E genotype. Neurology 50, 164–168 (1998). This study indicated that insulin levels can vary in the plasma and in the CSF, and it correlated these levels with severity of dementia in Alzheimer's disease.
Teede, H. J., Hutchison, S., Zoungas, S. & Meyer, C. Insulin resistance, the metabolic syndrome, diabetes, and cardiovascular disease risk in women with PCOS. Endocrine 30, 45–53 (2006).
Walker, D. W., McColl, G., Jenkins, N. L., Harris, J. & Lithgow, G. J. Evolution of lifespan in C. elegans. Nature 405, 296–297 (2000).
Golden, J. W. & Riddle, D. L. The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food and temperature. Dev. Biol. 102, 368–378 (1984).
Holzenberger, M. et al. Experimental IGF-I receptor deficiency generates a sexually dimorphic pattern of organ-specific growth deficits in mice, affecting fat tissue in particular. Endocrinology 142, 4469–4478 (2001).
Patel, D. S. et al. Clustering of genetically defined allele classes in the Caenorhabditis elegans DAF-2 insulin/IGF-1 receptor. Genetics 178, 931–946 (2008).
Leibson, C. L. et al. The risk of dementia among persons with diabetes mellitus: a population-based cohort study. Ann. NY Acad. Sci. 826, 422–427 (1997).
Ott, A. et al. Diabetes mellitus and the risk of dementia: The Rotterdam Study. Neurology 53, 1937–1942 (1999).
Dillin, A., Crawford, D. K. & Kenyon, C. Timing requirements for insulin/IGF-1 signaling in C. elegans. Science 298, 830–834 (2002).
McCay, C. M., Crowell, M. F. & Maynard, L. A. The effect of retarded growth upon the length of life span and upon the ultimate body size. 1935. Nutrition 5, 155–171; discussion 172 (1989).
Lin, S. J. et al. Calorie restriction extends Saccharomyces cerevisiae lifespan by increasing respiration. Nature 418, 344–348 (2002).
Klass, M. R. Aging in the nematode Caenorhabditis elegans: major biological and environmental factors influencing life span. Mech. Ageing Dev. 6, 413–429 (1977).
Partridge, L., Piper, M. D. & Mair, W. Dietary restriction in Drosophila. Mech. Ageing Dev. 126, 938–950 (2005).
Mair, W., Piper, M. D. & Partridge, L. Calories do not explain extension of life span by dietary restriction in Drosophila. PLoS Biol. 3, e223 (2005).
Panowski, S. H., Wolff, S., Aguilaniu, H., Durieux, J. & Dillin, A. PHA-4/Foxa mediates diet-restriction-induced longevity of C. elegans. Nature 447, 550–555 (2007).
Larsen, P. L., Albert, P. S. & Riddle, D. L. Genes that regulate both development and longevity in Caenorhabditis elegans. Genetics 139, 1567–1583 (1995).
Honda, Y. & Honda, S. The daf-2 gene network for longevity regulates oxidative stress resistance and Mn-superoxide dismutase gene expression in Caenorhabditis elegans. FASEB J. 13, 1385–1393 (1999).
Lithgow, G. J., White, T. M., Melov, S. & Johnson, T. E. Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proc. Natl Acad. Sci. USA 92, 7540–7544 (1995).
Pinkston, J. M., Garigan, D., Hansen, M. & Kenyon, C. Mutations that increase the life span of C. elegans inhibit tumor growth. Science 313, 971–975 (2006).
Pinkston-Gosse, J. & Kenyon, C. DAF-16/FOXO targets genes that regulate tumor growth in Caenorhabditis elegans. Nature Genet. 39, 1403–1409 (2007).
Hightower, L. E. Heat shock, stress proteins, chaperones, and proteotoxicity. Cell 66, 191–197 (1991).
Outeiro, T. F. et al. Sirtuin 2 inhibitors rescue α-synuclein-mediated toxicity in models of Parkinson's disease. Science 317, 516–519 (2007).
Bossy-Wetzel, E., Schwarzenbacher, R. & Lipton, S. A. Molecular pathways to neurodegeneration. Nature Med. 10 (Suppl.), S2–S9 (2004).
Muchowski, P. J. & Wacker, J. L. Modulation of neurodegeneration by molecular chaperones. Nature Rev. Neurosci. 6, 11–22 (2005).
Author information
Authors and Affiliations
Ethics declarations
Competing interests
Andrew Dillin is the founder of Prosteosis Therapeutics, Inc.
Glossary
- Youthfulness
-
The physical capabilities and conditions, such as agility, speed of crawling, rate of food intake and tissue integrity, that are typical of young but not old animals.
- Proteostasis
-
The favourable protein-homeostasis condition, in which the protein's composition, distribution, integrity, folding and protein–protein interactions are properly maintained, enabling optimal function at the cellular level.
- Protein folding
-
The post-translational process that a polypeptide has to complete in order to attain its optimal three-dimensional structure. Protein folding is often assisted by chaperones. Proteins that fail to achieve their correct folding are termed 'misfolded proteins'.
- Proteome
-
All of the functional proteins in a cell at a certain time point. The proteome can change as a function of age, stress conditions, et cetera.
- Amyloid-β
-
(Aβ). A highly aggregative peptide that is associated with and causative of AD.
- PolyQ
-
A stretch of glutamine repeats in a protein sequence. Abnormally long polyQ stretches are associated with various diseases, including HD.
- Protofibril
-
A small, soluble protein aggregate that is thought to be the cause of various neurodegenerative maladies.
- Long-term potentiation
-
(LTP). A long-lasting form of synaptic plasticity that results in an increase in the strength of synaptic transmission and is thought to be critical for memory function.
- Transcriptome
-
The gene network that is regulated by an activated transcription factor under certain conditions. Different transcriptomes can be regulated by the same transcription factor under distinct conditions.
- Protein clearance
-
A set of cellular activities aimed at identifying misfolded proteins, targeting them to specialized degradation complexes and mediating their digestion by specialized degradation machineries, such as proteasomes or lysosomes.
- RNA interference
-
(RNAi). A technique used to knock down the expression of a specific gene by introducing a double stranded RNA molecule that complements the gene of interest and triggers the degradation of the gene's mRNA molecules.
Rights and permissions
About this article
Cite this article
Cohen, E., Dillin, A. The insulin paradox: aging, proteotoxicity and neurodegeneration. Nat Rev Neurosci 9, 759–767 (2008). https://doi.org/10.1038/nrn2474
Published:
Issue Date:
DOI: https://doi.org/10.1038/nrn2474
This article is cited by
-
The neurobiology of insulin-like growth factor I: From neuroprotection to modulation of brain states
Molecular Psychiatry (2023)
-
Dietary restriction ameliorates TBI-induced phenotypes in Drosophila melanogaster
Scientific Reports (2022)
-
Protein clearance strategies for disease intervention
Journal of Neural Transmission (2022)
-
Insulin-like growth factor I sensitization rejuvenates sleep patterns in old mice
GeroScience (2022)
-
Targeting Insulin-Like Growth Factor-I in Management of Neurological Disorders
Neurotoxicity Research (2022)
