
Abstract
Accumulating evidence indicates that a sub-population of cancer cells with stem-like properties, termed tumor-initiating cells (T-ICs), exist in many different kinds of malignancies, which have a pivotal role in tumorigenesis, tumor progression, metastasis and post-treatment relapse. However, how the stem-like properties of T-ICs are regulated remains obscure. Our previous study showed that reduction of let-7 microRNA (miRNA) in breast tumor-initiating cells (BT-ICs) contributes to the maintenance of their self-renewal capacity and undifferentiated status. In this study we show the effect of mir-30 reduction on the stem-like features of BT-ICs. Similar to let-7, mir-30 is reduced in BT-ICs, and the protein level of Ubc9 (ubiquitin-conjugating enzyme 9) and ITGB3 (integrin β3), the target genes of mir-30, is markedly upregulated. Enforced constitutive expression of mir-30 in BT-ICs inhibits their self-renewal capacity by reducing Ubc9, and induces apoptosis through silencing ITGB3. On the contrary, blocking the miRNA with a specific antisense oligonucleotide (ASO) in differentiated breast cancer cells revived their self-renewal capacity. Furthermore, ectopic expression of mir-30 in BT-IC xenografts reduces tumorigenesis and lung metastasis in nonobese diabetic/severe combined immunodeficient mice, whereas blocking mir-30 expression enhances tumorigenesis and metastasis. Together, our data suggest mir-30 as one of the important miRNAs in regulating the stem-like features of T-ICs.
Similar content being viewed by others
Introduction
Accumulating evidence suggests that many cancers are maintained in a hierarchical organization of rare, slowly dividing tumor-initiating cells (T-ICs), precursor cells and differentiated tumor cells. It is hypothesized that T-ICs, also termed as cancer stem cells, are the cell origin of the tumor and responsible for tumorigenesis, tumor progression, metastasis and relapse (Al-Hajj, 2007; Dalerba and Clarke, 2007). T-ICs have been identified in a large panel of cancers (Gupta et al., 2009), including hematologic, brain, breast, prostate, liver, pancreatic and colon cancers. T-ICs resemble tissue stem cells in a variety of specific properties, termed ‘stemness’, including unlimited self-renewal, multidirectional differentiation potential and tumor formation capacity in vivo (Jordan et al., 2006). Unlimited self-renewal is an important hallmark of T-ICs, which allows the maintenance of the pool of T-ICs through asymmetric cell division, and is sustained by tightly coordinated molecular pathways. Therefore, revealing the molecular mechanisms that govern the stemness of T-ICs not only provides further insights into the biology of cancer, but also helps to develop novel therapeutic strategies targeting the refractory cancer stem cells (Cho and Clarke, 2008). However, T-ICs are rare in the tumor. To solve this problem, we enriched breast tumor-initiating cells (BT-ICs) in our previous study by consecutively passaging breast cancer cells in nonobese diabetic/severe combined immunodeficient (NOD/SCID) mice under the treatment with low-dose chemotherapy (Yu et al., 2007), and this provides a way of getting almost unlimited numbers of T-ICs.
MicroRNAs (miRNAs) are short 20–22 nucleotide RNA molecules that are negative regulators of gene expression in a variety of eukaryotic organisms. It is known that dysregulation of miRNA contributes to the initiation and progression of cancer (Chen, 2005; Croce and Calin, 2005). Recent reports of our research and other studies have highlighted the importance of miRNAs to maintain the stemness of T-ICs (Yu et al., 2007; Ji et al., 2009). In our previous study, we compared miRNA expression profiles in self-renewing BT-ICs and differentiated non-BT-ICs from a breast cancer line and primary breast cancers. We found that a panel of miRNAs, including let-7, is reduced in a panel BT-ICs from both SK-3rd line and primary patient tumors. Among them, the let-7 family emerges as the most consistently and significantly reduced miRNA, resulting in upregulation of HRAS and HMGA2, which promote self-renewal and block epithelial differentiation, respectively. Therefore, let-7 regulates multiple stem-like properties in BT-ICs by silencing more than one genes involved in self-renewal and differentiation (Yu et al., 2007). However, enforced expression of let-7 alone is not sufficient to completely block tumor formation and progression, whereas the effect of other differentially expressed miRNAs on T-ICs remains largely unclear.
In our present study, we extend our previous findings by showing the effect of mir-30 reduction on the stemness of BT-ICs. Similar to let-7, the expression of mir-30 is reduced in BT-ICs, and its target genes, Ubc9 (ubiquitin-conjugating enzyme 9), an E2-conjugating enzyme essential for sumoylation, and integrin β3 (ITGB3), are upregulated at protein level. Enforced constitutive expression of mir-30 in BT-ICs inhibits their self-renewal capacity by knocking down Ubc9 and induces apoptosis by inhibiting Ubc9 and ITGB3. Furthermore, transduction of mir-30 or Ubc9 small hairpin RNA (shRNA) in BT-ICs reduces their ability to form tumors and metastasize in NOD/SCID mice, whereas mir-30 antisense oligonucleotide (ASO) enhances tumorigenesis and metastasis of SKBR3 breast cancer cells with low metastasis potential.
Results
mir-30 expression and function are reduced in BT-ICs
Our previous study showed differential miRNA expression in BT-ICs and differentiated breast cancer cells (Yu et al., 2007). In this study, using miRNA microarray with an updated miRNA probe set (miRBase Sequence Database, Release 10.0), we found that mir-30 is reduced by approximately 30-fold in BT-ICs when compared with the differentiated breast cancer cells (Supplementary Table 1). We then further evaluated the expression of miRNAs from the mir-30 family using quantitative reverse transcription–PCR (qRT–PCR) with specific primers, and found that all members from mir-30 family, including mir-30d, mir-30a-5p, mir-30e-5p, mir-30b and mir-30c, are reduced in mammospheric SK-3rd cells, which were obtained by passaging SKBR3 breast cancer line in NOD/SCID mice under low-dose chemotherapy, when compared with their in vitro differentiated progeny, obtained by attaching mammospheric SK-3rd cells to the culture plates precoated with collagen IV (Supplementary Figure 1). Notably, mir-30e, the most commonly studied member of the family (the term ‘mir-30’ is used in the following text), is barely detectable in SK-3rd mammosphere cells, but is markedly enhanced by >20-fold in the differentiated SK-3rd cells (Figure 1a, P<0.01), which is not significantly different from that in the parental SKBR3 cells (Figure 1a, P>0.05).

mir-30 miRNA is reduced in mammospheric SK-3rd cells and BT-ICs obtained from primary breast tumors. (a) Quantitative RT–PCR amplified for mir-30e shows that mir-30e expression is higher in mammospheric (cultured in mammospheres) SK-3rd cells than in adherent SK-3rd cells and the parental SKBR3 line (&P<0.01 vs adherent SK-3rd or SKBR3). (b) Mammospheric SK-3rd cells infected with a lenti-mir-30e show elevated mir-30e expression (*P<0.001 vs untransduced cells). In contrast, mir-30e expression is reduced in differentiated SK-3rd cells transduced with lenti-mir-30 antagomir when compared with the untransduced cells (&P<0.01). (c) Primary BT-ICs enriched by mammosphere assays or lin-CD44+CD24− fluorescence-activated cell sorting (FACS) are reduced in mir-30e when compared with either adherent cells or tumor cells that were not CD44+CD24− (*P<0.001). (d) Transduction with lentivirus carrying mir-30 antagomir, but not with the empty lentivector, reduces mir-30e level in non-CD44+CD24− cells (&P<0.01 vs untransduced cells; *P<0.001 vs untransduced non-CD44+CD24− cells). (e) Transduction with lenti-mir-30, but not with lentivector, enhances mir-30 expression in primary mammospheres (*P<0.001 vs untransduced cells). Transduction with lentivirus carrying mir-30 antagomir reduces mir-30 level in the adherent primary cells (*P<0.001 vs untransduced cells). (f) Luciferase activity evaluated after transfection of a reporter vector carrying a mir-30 targeting sequence at its 3′-UTR is negligible in mammospheric SK-3rd, but increases upon differentiation or infection with lenti-mir-30 (*P<0.001 vs mammospheric SK-3rd). Infection with a lentivirus vector carrying mir-30 antagomir reduces endogenous or exogenous mir-30 activity (*P<0.001 vs untransfected cells). Error bars correspond to mean±s.d.
Infection of mamospheric SK-3rd cells with a lentiviral vector carrying a mir-30e pre-miRNA expression cassette (lenti-mir-30) increases mir-30 expression to the level comparable to differentiated SK-3rd cells (Figure 1b). In contrast, infection of the differentiated SK-3rd cells, which express considerable amount of mir-30, with a lentivirus encoding specific mir-30e antagomirs (lenti-mir-30-ASO), but not with a lenti-lin4-ASO, reduces the miRNA level by nearly threefold (Figure 1b). Moreover, lenti-mir-30-ASO results in reduction of all members of mir-30 to a similar extent (Supplementary Figure 1), suggesting a universal blocking effect of mir-30e-ASO to the mir-30 family.
To confirm that the results obtained from the SK-3rd line are physiologically relevant to primary breast cancers, we examined mir-30 expression in the BT-ICs isolated from primary breast cancers by qRT–PCR with a mir-30e-specific primer. Primary BT-ICs were enriched by mammosphere cultures (Ponti et al., 2005) or by sorting freshly isolated cells for lin-CD44+CD24− cells (Al-Hajj et al., 2003). Similar to SK-3rd cells, both the primary BT-ICs from mammospheres and sorted lin-CD44+CD24− cells are reduced in mir-30 when compared with either adherent cells or tumor cells that were not CD44+CD24− (Figure 1c, P<0.001). In addition, transduction with lenti-mir-30-ASO reduces mir-30 expression by >50% in adherent primary breast cancer cells or sorted non-CD44+CD24− cells (Figures 1d and e, P<0.01). On the other hand, infection with lenti-mir-30 increases the miRNA expression by approximately eight folds in primary mammospheres (Figure 1e, P<0.001). Taken together, these data indicated that mir-30 miRNAs are significantly reduced in mammospheric SK-3rd cells and BT-ICs obtained from primary patient tumors, whereas lentivirus expressing mir-30 pre-miRNA efficiently restores the expression of mir-30 in these cells.
To further investigate the targeting function of mir-30, we transfected a luciferase reporter vector containing a mir-30 target sequence cloned to its 3′ untranslated region (UTR) into mammospheric SK-3rd and differentiated SK-3rd, and evaluated the percentage of luciferase suppression relative to the cells transfected with a control luciferase reporter vector without mir-30 target sequence. Luciferase activity is suppressed by 65% in differentiated SK-3rd cells, whereas there is almost no suppression in mammospheric SK-3rd (Figure 1f, P<0.001). Infection of mammospheric SK-3rd with a lenti-mir-30 significantly restores the suppression of luciferase activity by approximately 47% (Figure 1f, P<0.001). Co-transfecting the cells with both the reporter vector and the mir-30 ASO significantly relieves the suppression in luciferase activity in adherent SK-3rd with abundant endogenous mir-30 and in mammospheric SK-3rd with ectopic mir-30 expression (Figure 1f, P<0.001). Hence, miR-30 targeting function is reduced in BT-ICs, which can be restored by transduction of its precursor expression cassette.
Ubc9 and ITGB3 serve as target genes for mir-30
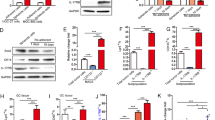
Ubc9, an E2-conjugating enzyme for sumoylation, has been reported as a target gene for mir-30. In addition, with the computer-based sequence analysis software TargetScan, we found that ITGB3 (CD61) 3′UTR mRNA has a perfect match with the seed sequence of mir-30 (Supplementary Table 2). Using qRT–PCR assay, we found that mammospheric SK-3rd cells abundantly express ITGB3 mRNA, which is nearly seven folds higher than the differentiated SK-3rd cells (P<0.01), and three folds higher than the parental SKBR3 cells (Figure 2a, P<0.05). In contrast, Ubc9 mRNA expression is comparable in mammospheric and adherent SK-3rd, which is not significantly different from the parental SKBR3 cells (Figure 2b, P>0.05). In agreement, the ITGB3 mRNA is significantly higher in BT-ICs enriched from primary tumors of breast cancer patients by mammosphere culture or by CD44+CD24− sorting than that in the non-enriched cancer cells or the non-CD44+CD24− cancer cells (Figure 2a, P<0.01), whereas Ubc9 mRNA is not significantly different among these cells (Figure 2b, P>0.05). Furthermore, immunoblotting with anti-ITGB3 and anti-Ubc9 antibodies showed both the ITGB3 and Ubc9 proteins are significantly elevated in mammospheric SK-3rd cells compared with the differentiated SK-3rd cells and the parental SKBR3 cells (Figures 2c and d).

ITGB3 and Ubc9 serve as direct targets genes for mir-30. (a) qRT–PCR assay shows that mammospheric SK-3rd has a higher level of ITGB3 mRNA than the differentiated SK-3rd cells (&P<0.01) and parental SKBR3 cells (#P<0.05; left panel). The mRNA level of ITGB3 in mammospheric SK-3rd cells transfected with lenti-mir-30 is not changed (P>0.05; left panel). ITGB3 mRNA level is higher in BT-ICs, enriched from primary breast tumors by mammosphere culturing or CD44+CD24− sorting, than non-enriched cancer cells or non-CD44+CD24− cancer cells (&P<0.01; right panel). (b) On the other hand, qRT–PCR assay shows that Ubc9 mRNA expression was comparable in mammospheric and adherent SK-3rd, which was not significantly different from parental SKBR3 cells (P>0.05; left panel). The mRNA level of Ubc9 in mammospheric SK-3rd cells transfected with lenti-mir-30 is not changed (P>0.05; left panel). Ubc9 mRNA is not different between mammosphere-enriched primary breast cancer cells and non-enriched cancer cells, or between CD44+CD24− sorted primary breast cancer cells and non-CD44+CD24− cancer cells(P>0.05) (right panel). (c, d) Immunoblotting with anti-ITGB3 and anti-Ubc-9 antibodies shows that ITGB3 (c) and Ubc-9 (d) proteins are elevated in mammospheric SK-3rd cells compared with differentiated SK-3rd cells and parental SKBR3, and are knocked down to the level comparable to their differentiated progenies by infection with lenti-mir-30e. (e, f) Transfection of mammospheric SK-3rd cells with a mir-30e mimics suppresses the luciferase activity of a co-transfected reporter vector containing the mir-30 recognition sequence cloned from the 3′UTR of the ITGB3 mRNA (luc-ITGB3-3′UTR) (e, left panel) and Ubc-9 mRNA (luc-Ubc9-3′UTR) (f, left panel), but has no effect on the activity of a co-transfected reporter vector containing a mutant 3′UTR of the ITGB3 mRNA (luc-mut-ITGB3-3′UTR) (e, left panel) and the Ubc9 mRNA (luc-mut-Ubc9-3′UTR) (f, left panel) (*P<0.001 vs miRNA mimic control). Vice versa, transfection of the adherent SK-3rd cells with the miR-30 inhibitor increased the luciferase activity of a co-transfected luc-ITGB3-3′UTR (e, right panel) and luc-Ubc9-3′UTR (f, right panel), but has no effect on the activity of a co-transfected luc-mut-ITGB3-3′UTR (e, right panel) and luc-mut-Ubc9-3′UTR (f, right panel) (&P<0.01 vs inhibitor control). Error bars correspond to mean±s.d.
To examine whether mir-30 affects ITGB3 and Ubc9 expression by targeting the 3′UTR, we infected the mammospheric SK-3rd cells with lenti-mir-30, and monitored the mRNA expression of ITGB3 and Ubc9 with qRT–PCR and their protein levels with western blotting. Although the mRNA levels of ITGB3 and Ubc9 in mammospheric SK-3rd cells are almost not changed after transduction with lenti-mir-30 (Figures 2a and b), ITGB3 and Ubc9 proteins in these cells are significantly knocked down to the level comparable to their differentiated progenies (Figures 2c and d). The effect is specific as ITGB3 and Ubc9 proteins in the untransduced cells or the cells transduced with an empty vector remain unchanged (Figures 2c and d). On the basis of these observations, Ubc9 and ITGB3 are potential targets for miR-30.
The targeting function of mir-30 to the 3′UTR of ITGB3 and Ubc9 mRNA was further elucidated using luciferase reporter constructs containing the mir-30 recognition site or a mutated sequence at this site cloned from the 3′UTR of the ITGB3 mRNA or the Ubc9 mRNA immediately downstream of luciferase gene. As shown in Figures 2e and f, transfection of mammospheric SK-3rd cells with a mir-30 mimic suppresses the luciferase activity of the reporter vectors containing the 3′UTR of a wild-type ITGB3 or a wild-type Ubc9, whereas mir-30-ASO increases that of the adherent SK-3rd. However, both mir-30 and mir-30 ASO had no effect on the luciferase activity of the reporters with a mutant ITGB3 or a mutant Ubc9 3′UTR. Therefore, miR-30 regulates ITGB3 and Ubc9 expression by directly targeting their 3′UTR and inducing translational repression. Collectively, our data insinuate that mir-30 reduction in BT-ICs may result in Ubc9 and ITGB3 protein elevation by relieving translational repression at the 3′UTR of the target genes, whereas other mechanisms at transcriptional level may also contribute causally to the upregulation of ITGB3 mRNA in BT-ICs.
mir-30 reduction contributes to maintain self-renewal of BT-ICs
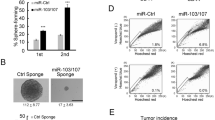
To investigate whether mir-30 reduction has a role in maintaining the population of BT-ICs, we first studied the effect of ectopic mir-30 expression on the self-renewal of SK-3rd by examining the percentage and size of mammospheres formed by SK-3rd cells that had been transduced with lenti-mir-30 or lenti-mir-30-ASO. As a positive control, ectopic let-7 expression was used. We found that mammospheric SK-3rd cells infected with lenti-mir-30 or lenti-let-7 form approximately 65 and 87% less mammospheres, respectively, than the uninfected SK-3rd or SK-3rd cells infected with an empty lentiviral vector or a lentiviral vector expressing an irrelevant lin4 miRNA (lenti-lin4; P<0.0001, Figure 3a). Furthermore, infection with lenti-mir-30 and lenti-let-7 synergistically reduces the percentage of mammospheres formed by mammospheric SK-3rd (P<0.0001, Figure 3a), suggesting that the two miRNAs may target to different pathways that are involved in regulating the self-renewal capacity of BT-ICs. Similarly, transduction of mammospheric SK-3rd with lenti-mir-30 or lenti-let-7, but not with lenti-lin4, significantly reduces the number of cells per mammosphere after 15 days in suspension culture, and transduction of both miRNAs synergistically reduces the cell number in the mammosphere (Figure 3c). Conversely, infection of the differentiated SK-3rd cells with lenti-mir-30-ASO, instead of lenti-lin4-ASO, increases their ability to form mammospheres by nearly three folds (Figure 3e).

Ectopic expression of mir-30e reduces mammosphere formation and enhances apoptosis in BT-ICs obtained from SK-3rd and primary breast cancers. (a, b) Suspension cultures of dissociated SK-3rd cells (a) or primary breast cancer cells sorted for lin–CD44+CD24−/low phenotype (b), infected with lenti-mir-30 or lenti-let7, instead of lenti-lin4 or an empty vector, form fewer mammospheres ($P<0.0001 vs untransfected control). Infection of SK-3rd cells (a) or primary breast cancer cells sorted for lin–CD44+CD24−/low phenotype (b) with both lenti-mir-30 and lenti-let-7 synergistically reduced the percentage of mammospheres ($P<0.0001 vs untransfected control or mir-30 transfected cells; *P<0.001 vs let-7 transfected cells). Besides, SK-3rd cells (a) or primary breast cancer cells sorted for lin–CD44–CD24−/low phenotype (b), infected with lenti-Ubc9-shRNA, but not lenti-GFP-shRNA, form fewer mammospheres ($P<0.0001 vs untransfected control). (c, d) The cell number per mammosphere formed by SK-3rd cells (c) or primary breast cancer cells sorted for lin–CD44+CD24−/low phenotype (d), infected with lenti-mir-30 or lenti-let7, but not with lenti-lin4 or an empty vector, is significantly reduced ($P<0.0001 vs untransfected control). Infection of SK-3rd cells (c) or primary breast cancer cells sorted for lin–CD44+CD24−/low phenotype (d) with both lenti-mir-30 and lenti-let-7 synergistically reduced the cell number per mammosphere ($P<0.0001 vs untransfected control or mir-30e transfected cells; *P<0.001 vs let-7 transfected cells). Besides, SK-3rd cells (c) or primary breast cancer cells sorted for lin–CD44+CD24−/low phenotype (d), infected with lenti-Ubc9-shRNA, but not lenti-GFP-shRNA, generate mammospheres with fewer cell number ($P<0.0001 vs untransfected control). (e, f) Conversely, differentiated SK-3rd cells (e) non-CD44+CD24− breast cancer cells (f) transduced with mir-30 ASO, but not with lin-4 ASO, generate more mammospheres ($P<0.0001 vs untransfected control). (g) Phase-contrast microscopy shows that transduction of SK-3rd cells with lenti-mir-30, lenti-ITGB3-shRNA or lenti-Ubc9-shRNA, instead of lenti-lin4, lenti-GFP or an empty vector, reduces sphere formation in anoikis assay. (h) Transduction of the SK-3rd cells with lenti-mir-30e, lenti-ITGB3 or lenti-Ubc9, but not with lenti-lin4, lenti-GFP-shRNA or an empty vector, increases the percentage of Annexin V (h) and TdT-mediated dUTP nick end labeling (TUNEL) (i) positive cells ($P<0.0001 compared with untransduced cells).
Furthermore, CD44+CD24− cells that were isolated from primary breast cancers and infected with lenti-mir-30 or lenti-let-7, instead of lenti-lin4, give rise to significantly fewer mammospheres (Figure 3b, P<0.001), and infection with both lenti-mir-30 and lenti-let-7 further decreases the capacity of mammosphere formation (Figure 3b, P<0.0001 vs lenti-let-7). Moreover, the number of cells per mammosphere formed by CD44+CD24− breast cancer cells infected with lenti-mir-30 or lenti-let-7 decreases by more than 50 and 70%, respectively, compared with empty vector and lenti-lin4 (Figure 3d, P<0.0001). On the other hand, transducing non-CD44+CD24− breast cancer cells with lenti-mir-30-ASO, instead of lenti-lin4-ASO, increases their ability to form mammospheres by more than two folds (Figure 3f, P<0.0001). In agreement, the percentage of CD44+CD24− cells is dramatically increased by transduction with lenti-mir-30-ASO, but not with lenti-lin4-ASO or an empty vector, in unsorted primary breast cancer cells (Supplementary Figure 2, P<0.001). Therefore, our data in breast cancer line and primary breast cancers suggest that enforced mir-30 expression in BT-ICs inhibits their self-renewing capacity as let-7 does, and both miRNAs may act synergistically on BT-ICs.
To further explore whether the enhanced capability of mammosphere formation induced by mir-30 loss is mediated by elevation of ITGB3 and Ubc9, we silenced ITGB3 and Ubc9 expression by specific RNA interference. Interestingly, transduction of ITGB3-shRNA with lentivirus (lenti-ITGB3-shRNA) in mammospheric SK-3rd cells reduces the formation of secondary mammospheres by only approximately 7.3% (Supplementary Figure 3A, P>0.05) and the size of spheres by only 11% after 15 days of suspension culture (Supplementary Figure 3C, P>0.05) when compared with the empty vector or lenti-green fluorescence protein (GFP)-shRNA. This finding suggests that silencing ITGB3 is obviously less potent to inhibit self-renewal of BT-ICs than ectopic mir-30 expression. In contrast, transduction with lenti-Ubc9-shRNA reduces self-renewal capacity of mammospheric SK-3rd by approximately 32% (Figure 3a, P<0.0001 vs lenti-GFP-shRNA), although not to the extent of lenti-mir-30 transduction. Similarly, secondary mammosphere formation and the number of cells per sphere in the sorted CD44+CD24− primary breast cancers were significantly reduced by silencing Ubc9 (Figures 3b and d, P<0.001 vs lenti-GFP-shRNA), but not by silencing ITGB3 (Supplementary Figures 3B and D, P>0.05 vs lenti-GFP-shRNA). These data suggest that mir-30 reduction results in enhanced self-renewal of BT-ICs partly through Ubc9, instead of ITGB3, upregulation.
mir-30 reduction inhibits apoptosis in BT-ICs through ITGB3 and Ubc9 upregulation
Anoikis is a form of programmed cell death (apoptosis) that is induced when anchorage-dependent cells are detached from the surrounding extracellular matrix (Gilmore, 2005). However, T-ICs are believed to be anoikis resistant (Glinsky et al., 2005) and they have the capability of ‘anchorage-independent’ or ‘spheroid’ growth that allows them to escape from extracellular matrix attachment and develop metastasis. To show whether mir-30 and its target genes have a role in anoikis resistance and apoptosis, we used anoikis assay and apoptosis detection by TUNEL (TdT-mediated dUTP nick end labeling) and Annexin V staining. Interestingly, the percentage of SK-3rd cells with the capacity of anchorage-independent growth is dramatically reduced by approximately eight folds after transduction with lenti-mir-30 or lenti-ITGB3-shRNA or lenti-Ubc9-shRNA, compared with the untransduced SK-3rd cells or cells transduced with empty vector or lenti-GFP-shRNA (Figure 3g and Supplementary Figure 4, P<0.0001). Moreover, transduction of mammospheric SK-3rd cells with lenti-mir-30 or lenti-ITGB3-shRNA or lenti-Ubc9-shRNA also tremendously increases the proportion of TUNEL-positive cells (Figure 3h, P<0.0001) and enhances the percentage of Annexin V-positive cells (Figure 3i, P<0.0001). These results suggest that reduction of mir-30 is responsible for anoikis resistance and apoptosis inhibition in BT-ICs through upregulating ITGB3 and Ubc9.
Ectopic miR-30 expression reduces the tumorigenesis and metastasis of BT-ICs
A major feature defining T-ICs is their strong ability to establish themselves as tumor xenograft (Dalerba et al., 2007). Thus, we inoculated various number of mamospheric SK-3rd cells that were transduced with lenti-mir-30, lenti-Ubc9-shRNA or lenti-ITGB3-shRNA into the fat pads of NOD/SCID mice to generate breast tumors, and investigated the influence of mir-30 and its target genes in tumorigenesis and metastasis of BT-ICs. As shown in Table 1, all the 10 mice xenografted with 1 × 104 mamospheric SK-3rd cells that were untransduced or transduced with empty vector or lenti-lin4 generate tumors from days 33 to 36 after inoculation. Furthermore, within 14 days after tumors are identified, their sizes reach 1.8±0.7 cm in diameter. In contrast, only 5 out of 10 mice (50%) xenografted with mamospheric SK-3rd cells transduced with lenti-mir-30 and 6 out of 10 mice with lenti-Ubc9-shRNA generate tumors, and it takes 17–24 days for the palpable tumors to reach 2.0 cm in diameter (Figure 4a). In addition, when 2 × 106 mamospheric SK-3rd cells were inoculated, transduction with lenti-mir-30 or lenti-ITGB3-shRNA also retards tumor growth, but the effect of lenti-ITGB3-shRNA is weaker (Supplementary Figure 5A; Supplementary Table 3). Moreover, the average tumor weight of mir-30-expressing SK-3rd xenografts, as well as Ubc9- or ITGB3-silenced SK-3rd xenografts, is dramatically reduced compared with that of untransduced SK-3rd xenografts (Supplementary Figure 6A, P<0.001; Supplementary Figure 5B, P<0.0001). However, transduction with lenti-lin4, lenti-GFP-shRNA or empty vector does not significantly influence the tumor weight of SK-3rd xenografts (Supplementary Figure 6B, P>0.05; Supplementary Figure 5B, P>0.05). These data suggest that mir-30 and its targets Ubc9 and ITGB3 have an important role in tumorigenesis of BT-ICs. In addition, hematoxylin and eosin staining shows that the overexpression of mir-30 or silencing of Ubc9 or ITGB3 with lentivirus vector does not change the histological features of the tumors formed by mamospheric SK-3rd cells (Figure 4c, Supplementary Figure 5C).

Enforcing mi-30 expression reduces tumorigenesis and metastasis of mammospheric SK-3rd, whereas inhibition of miR-30 enhances tumorigenesis and metastasis of SKBR3. Mammospheric SK-3rd cells or SKBR3 cells were inoculated into the mammary fat pads of NOD/SCID mice at 1 × 104 or 1 × 106 cells, respectively, and tumor volume was followed until the animals were killed on days 36 or 51 after implantation, respectively. (a) Mammospheric SK-3rd cells infected with lenti-mir-30 or lenti-Ubc9-shRNA, but not with the empty lentivirus vector or with lenti-lin4 or lenti-GFP-shRNA, generate fewer tumors, which grow more slowly when compared with the uninfected controls. (b) Quantitative RT–PCR using a human HRPT-specific primer shows that hHRPT expression in mouse lungs was markedly reduced in the mice xenografted with mammospheric SK-3rd cells infected with lenti-mir-30 or lenti-Ubc9-shRNA, but not with the empty lentivirus vector or lenti-lin4 or lenti-GFP-shRNA when compared with the uninfected controls (&P<0.01; #P<0.05 compared with untransduced cells). Error bars correspond to mean±s.d. (c) Hematoxylin and eosin staining (H&E, × 200) for tissue sections of the tumor xenografts (upper panel) and the lungs (lower panel) of the mice xenografted with uninfected mammospheric SK-3rd cells or cells infected with the empty lentivirus vector, or with lenti-lin4, lenti-mir-30, lenti-Ubc9-shRNA or lenti-GFP-shRNA, had similar histology by H&E staining (magnification). (d) SKBR3 cells infected with lenti-mir-30-ASO, but not with the empty lentivirus vector or with lenti-lin4-ASO, generate more tumors, which grow faster when compared with the uninfected controls. (e) Quantitative RT–PCR using a human HRPT-specific primer shows that hHRPT expression in mouse lungs was markedly increased in the mice xenografted with SKBR3 cells infected with lenti-mir-30-ASO, but not with the empty lentivirus vector or lenti-lin4-ASO when compared with the uninfected controls (&P<0.01 vs untransfected cells). Error bars correspond to mean±s.d. (f) H&E staining ( × 200) for tissue sections of the tumor xenografts (upper panel) and the lungs (lower panel) of the mice xenografted with uninfected SKBR3 cells or cells infected with the empty lentivirus vector, or with lenti-lin4-ASO or lenti-mir-30-ASO, had similar histology by H&E staining (magnification).
Furthermore, the ability to metastasis is also an important feature of T-ICs (Dalerba and Clarke, 2007). To evaluate the influence of miR-30 expression on the metastasis of BT-ICs, mice were killed when the tumor diameter reached 2 cm and lung metastases were examined with microscopy. Massive lung metastases were visualized under microscopy with hematoxylin and eosin staining in 9 out of 10 mice inoculated with untransduced mamospheric SK-3rd cells, whereas only 3 of the 10 mice injected with mir-30-expressing or Ubc-9-silenced SK-3rd cells, and only 4 of the 10 mice injected with ITGB3-silenced SK-3rd cells developed microscopically observable lung metastasis upon killing (Table 1; Figure 4c; Supplementary Figure 5C; Supplementary Table 3). However, transduction with empty viral vector, lenti-lin4 or lenti-GFP-shRNA does not influence microscopically observable lung metastasis (Figure 4c; Supplementary Figure 5C). This is in agreement with the measurement of wet lung weight, as the average lung weight of the mice xenografted with mir-30-expressing or Ubc9- or ITGB3-silenced mamospheric SK-3rd cells is significantly lower than that of the mice injected with control SK-3rd cells (Supplementary Figure 6A, P<0.001; Supplementary Figure 5D, P<0.01). Moreover, hematoxylin and eosin staining reveals that the lesions of lung metastasis in the mice xenografted with lenti-mir-30 or lenti-Ubc9-shRNA or lenti-ITGB3-shRNA transduced mamospheric SK-3rd cells are barely observed or dispersed among alveoli, whereas the lung metastasis lesions formed by SK-3rd cells transduced by empty vector or lenti-lin4 or lenti-GFP-shRNA are massive and obvious (Figure 4c; Supplementary Figure 5C). This is in agreement with the quantitative data, as a significant reduction of the human HPRT mRNA in the mouse lungs with tumor metastasis was observed in the mice xenografted with mammospheric SK-3rd cells infected with lenti-mir-30 (n=3, Figure 4b, P<0.01) or lenti-Ubc9-shRNA (n=3, Figure 4b, P<0.05) or lenti-ITGB3-shRNA (n=4, Supplementary Figure 5E, P<0.01) when compared with that of the mice inoculated with cells transduced with the empty vector.
On the other hand, we transduced parental SKBR3 cells that were low metastatic and expressed high levels of mir-30 with lenti-mir30-ASO, and evaluated tumor formation and metastasis in NOD/SCID mice after inoculation of 1 × 106 cells into the fat pads. In contrast to enforced mir-30 expression, transduction with lenti-mir30-ASO, but not with lenti-lin4-ASO, accelerates tumor formation (Figure 4d), increases the number of mice with lung metastasis (Table 2, P<0.001), increases tumor weight and lung weight (Supplementary Figure 6B, P<0.001; P<0.01) and enhances hHPRT mRNA expression in mouse lungs (Figure 4e, P<0.01) when compared with transduction with empty vectors. Moreover, lenti-mir30-ASO transduction does not alter the histological structure of the xenografts, but enhances microscopic lung metastasis (Figure 4f). Collectively, these data indicate that mir-30 reduction in BT-ICs promotes tumorigenesis and lung metastasis of the xenografted cancers by elevating Ubc9 and ITGB3.
Discussion
In this study we used SK-3rd cells, a previously established breast cancer line enriched for T-ICs, as our research model to evaluate the role of mir-30 microRNA in regulating the biological features of BT-ICs. We found that mir-30 is markedly reduced in SK-3rd mammospheres as well as in primary BT-ICs from breast cancer patients. Introduction of mir-30 into BT-ICs not only reduces their ability of self-renewal in vitro and in vivo, but also inhibits their capacity of anoikis resistance and increased apoptosis. Ubc9 (Wu et al., 2009) and ITGB3 are two direct target genes of mir-30, and they are both upregulated in BT-ICs because of mir-30 reduction. Furthermore, Ubc9 elevation maintains the self-renewing capacity of BT-IC, whereas overexpression of both Ubc9 and ITGB3 contributes to the anoikis resistance and apoptosis inhibition in the T-ICs.
We previously showed that the miRNA profile in mammosphere-derived SK-3rd cells is different from their differentiated progenies (Yu et al., 2007), and microarray data showed that a number of miRNAs, including let-7, miR-15, miR-16, miR-200 and miR-30, are markedly reduced in BT-ICs. Along with let-7, in this study we confirmed that mir-30 is barely detected in T-ICs originated from breast cancer line and primary breast cancers. However, we previously showed that not all miRNAs that are reduced in BT-ICs contribute to the ‘stemness’ of the initiating cells, as introduction of miR-15 and miR-16 did not influence self-renewal of BT-ICs (Yu et al., 2007). In the present study, in line with the findings of let-7, mir-30 reduction helps to maintain the self-renewal capability of BT-ICs, whereas ectopic expression of mir-30 in BT-ICs markedly reduces mammosphere formation in vitro and tumorigenesis in vivo, although the effect of mir-30 is not as prominent as let-7. On the contrary, blocking mir-30 with antagomir enhances tumorigenesis. These findings corroborate our previous observation that let-7 alone is not sufficient to completely abolish the self-renewing ability and tumorigenesis of BT-IC. More importantly, in this study we show that introduction of both let-7 and mir-30 lends a more complete inhibition to self-renewal of BT-IC when compared with either miRNA alone, and the rate of mammosphere formation in BT-IC with ectopic expression of both let-7 and mir-30 is comparable to non-BT-IC. The synergistic effect of let-7 and mir-30 on self-renewal inhibition can be ascribed to the distinct target molecules of the two miRNAs that are involved in different signaling pathways regulating the self-renewing capacity of T-ICs. As let-7 reduction in BT-ICs results in upregulation of HRAS and HMGA2, which promote self-renewal and block epithelial differentiation, respectively (Yu et al., 2007), mir-30 reduction leads to upregulation of Ubc9 and ITGB3, which enhance self-renewal and inhibit apoptosis. Hence, dysregulation of more than one miRNA results in aberrant expression of multiple oncogenes and tumor-suppressing genes, and ultimately regulates the biology of T-ICs. Therefore, a combination of tumor-initiating miRNA mimics might be necessary to eradicate cancer-initiating cells for a more efficient therapeutic approach against solid malignant tumors.
As mir-30 is markedly reduced in BT-ICs, expression of Ubc9 is upregulated at protein instead of mRNA level. This is in line with a previous report suggesting that Ubc9 is a direct target of mir-30, and the latter regulates Ubc9 expression mainly through translation repression (Wu et al., 2009). We confirm these findings by assays with a luciferase reporter carrying the Ubc9 3′-UTR, which indicates that mir-30 directly suppresses the luciferase activity of the vector with wild-type Ubc9 3′-UTR but not the one with mutated sequence at the mir-30e target site. Moreover, Ubc9 protein, instead of mRNA, is elevated in BT-ICs, and introduction of mir-30 greatly reduces its protein expression but does not influence the level of its mRNA, insinuating that dysregulation of Ubc9 in T-ICs is mainly through translation repression by mir-30. Ubc9 is an E2-conjugating enzyme essential for protein sumoylation, which modifies substrate proteins post-translationally by covalently attaching them with the small ubiquitin-related polypeptide SUMO (small ubiquitin-like modifier), and thus has a key role in nuclear trafficking, transcriptional regulation and stability maintenance of numerous proteins (Muller et al., 2001). The expression of Ubc9 protein is higher in breast cancers when compared with normal breast tissues, and ectopic expression of Ubc9 enhances, whereas suppression of Ubc9 function by the dominant-negative Ubc9 inhibits, tumor growth in the animal model, suggesting that Ubc9 has a causal role in tumorigenesis (Mo et al., 2005). In this study, we provide a further insight for the role of Ubc9 in tumorigenesis by showing that Ubc9 is more elevated in T-ICs cells than non-initiating cells, and silencing Ubc9 by RNA interference inhibits the self-renewal ability and tumorigenesis of BT-ICs. This is in line with the findings that Ubc9 is upregulated in neural stem cells, embryonic stem cells and embryonic cancer cells, and maintains the proliferation of these cells (Park et al., 2007; Zhang et al., 2007; Watanabe et al., 2008). Therefore, Ubc9 expression tends to increase in the cells with enhanced self-renewal ability, and regulates the stem-like features of the cells. The role of Ubc9 in self-renewal and tumorigenesis of T-ICs is likely because of the fact that Ubc9 regulates the expression of numerous proteins that are related to the self-renewing process by sumoylation. Among them, sumoylation of oct4 induced by Ubc9 results in its overexpression in embryonic stem cells, and thus contributes to the self-renewal and pluripotency of the cells (Park et al., 2007). We have shown that oct4 protein is upregulated in BT-ICs and decreases when the cells differentiate (Yu et al., 2007). Therefore, Ubc9 overexpression induced by mir-30 reduction may account for the abundance of oct4 in BT-IC and its subsequent reduction after differentiation.
In addition to Ubc9, we found that ITGB3 is also a direct target of mir-30, as mir-30 suppresses the luciferase activity of a reporter carrying the sequence of the ITGB3 3′UTR, whereas mutation at this target sequence abolishes the suppression effect. Moreover, introduction of mir-30 into BT-ICs specifically suppresses ITGB3 expression at protein but not mRNA level, suggesting that mir-30 also inhibits ITGB3 through translation repression. However, unlike Ubc9 with only protein overexpression, ITGB3 is elevated in BT-ICs at both mRNA and protein levels, insinuating that other regulatory mechanisms at transcription level may also be involved in ITGB3 upregulation in BT-ICs. In normal breast epithelia, however, ITGB1, rather than ITGB3, is upregulated in mammary stem cells, whereas ITGB3 is overexpressed in luminal progenitors (Pontier and Muller, 2009). The discrepancy of different integrin subtypes in normal and malignant mammary stem cells may insinuate the difference in biological features between normal and malignant stem cells, and may be further used as cell surface markers to distinguish these cells. Functionally, we show that ITGB3 upregulation is important for anoikis resistance of BT-ICs, and inhibiting ITGB3 expression with RNA interference leads to massive apoptosis of the cells. Anoikis resistance is believed to be a key property of T-ICs so that they can migrate and develop metastasis in distal organs (Glinsky et al., 2005). Integrins expressed on cancer cell surface sense the mechanical stimuli arising from the matrix, and thereby convert these stimuli to downstream signals to modulate cell viability (Zhan et al., 2004). Among various subtypes of integrins, ITGB3 is potent in mediating attachment of epithelial cells to the surrounding matrix fibers and thus transducing surviving signals to the adherent cells (Mizejewski, 1999).
Other than inhibiting tumorigenesis, we show that enforced mir-30 expression or silencing Ubc9 or ITGB3 expression with RNA interference in BT-IC xenografts significantly inhibits lung metastasis, whereas blocking mir-30 with antagomirs in breast cancer line with low metastatic capability enhances the metastasis. This is in agreement with previous reports that both Ubc9 and ITGB3 contribute to promote the invasiveness and metastasis of breast cancers. Therefore, the metastasis inhibitory effect of mir-30 is probably related to its targets, Ubc9 and ITGB3. Introduction of mir-30 reduces the number of BT-ICs with high metastatic potential by inhibiting self-renewal of the cells through Ubc9, and induces anoikis of BT-ICs through ITGB3. Both of these machineries may contribute to inhibit metastasis that is raised by BT-ICs.
In summary, this study highlights the importance of mir-30 for the stemness of T-ICs. Reduction in mir-30 maintains self-renewal and tumorigenic capability of BT-ICs through upregulating Ubc9, and inhibits apoptosis of BT-ICs through both Ubc9 and ITGB3. In align with our previous study, these data again suggest that tumor-inhibiting miRNA molecules may serve as therapeutic agents for breast cancer-targeting T-ICs. To achieve this goal, further studies are needed to develop methods to efficiently deliver miRNA mimics into BT-ICs.
Materials and methods
Detailed procedures are provided in the Supplementary Data.
Breast tumor-initiating cell line
The breast tumor-initiating cell line SK-3rd used in this study were previously established by consecutively passaging breast cancer cell line SKBR3 in NOD/SCID mice under the pressure of chemotherapy (Yu et al., 2007).
Mammosphere culture
SK-3rd and SKBR3 cells were cultured in suspension at a clonal density of 1000 cells/ml in serum-free Dulbecco's modied Eagle's medium/F12 (BioWhittaker, Walkersville, MD, USA), supplemented with B27 (1:50, Invitrogen, Carlsbad, CA, USA), 20 ng/ml epidermal growth factor (BD Biosciences, San Jose, CA, USA), 0.4% bovine serum albumin (Sigma, St Louis, MO, USA) and 4 μg/ml insulin (Sigma), as previously described (Dontu et al., 2003; Yu et al., 2007; see details in Supplementary Experimental Procedures).
miRNA microarray analysis
The method for microRNA microarray analysis was described in our previous report (Yu et al., 2007) using an updated miRNA probe set (the NCode miRNA Microarray Probe Set V3; Invitrogen).
Quantitative RT–PCR
Real-time RT–PCR was performed using an ABI Prism 7900 Sequence Detection System (Perkin-Elmer Applied Biosystems, Norwalk, CT, USA). SYBR green (Molecular Probes, Eugene, OR, USA) was used to detect PCR products (see details in Supplementary Experimental Procedures).
Primary tumor specimens
Tumors were obtained following a protocol approved by the ethics committee of the No. 2 Affiliated Hospital of Sun-Yat-Sen University from six consented female patients (median age 53 years) with biopsy-diagnosed poorly differentiated invasive ductal carcinomas of the breast. Tumors were mechanically disaggregated immediately and digested with collagenase as described (Yu et al., 2007). Single cancer cells were obtained by filtration through a 30 u filter (see details in Supplementary Experimental Procedures).
Flow cytometry
For cell surface staining, unfixed cells were incubated with fluorescein isothiocyanate-labeled anti-CD44 and phycoerythrin-labeled anti-CD24 (PharMingen, San Diego, CA, USA) at 4 °C. For staining cytoplasmic antigens, cells were permeabilized with the Caltag Laboratories (Burlingame, CA, USA) Fix and Perm kit and stained with fluorescein isothiocyanate-labeled CK14 and phycoerythrin-labeled CK18 (Neomarkers, Fremont, CA, USA). Cells were analyzed by flow cytometry on a FACScalibur instrument using CellQuest software (BD).
Transduction with lentivirus vectors
SK-3rd cells dissociated from the primary mammospheres were spin-infected with 1 ml of lentiviral supernatant containing 8 μg/ml polybrene for 2 h at a multiplicity of infection of 1:5, followed by incubation for 2 h at 37 °C. Transduction efficiency, evaluated by GFP expression, was >90%.
Mir-30 luciferase assay
To evaluate the miRNA function of mir-30, a pMIR-REPORTTM luciferase reporter vector with a mir-30 target sequence cloned into its 3′UTR (luc-mir-30-ts) (Ambion, Austin, TX, USA) was used (see details in Supplementary Experimental Procedures).
Western blot
Protein extracts were resolved through 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes, probed with rabbit polyclonal antibodies against ITGB3 (Abcam, Cambridge, MA, USA) or Ubc9 (Abcam), then with peroxidase-conjugated goat anti-rabbit Ig secondary antibody (Oncogene Research Product, Cambridge, MA, USA) and then visualized using chemiluminescence (Amersham, Arlington Heights, IL, USA).
Tumor implantation
Indicated numbers of SK-3rd cells dissociated from mammospheres or adherent parental SKBR3 cells were injected subcutaneously into the mammary fat pad of 5-week-old NOD/SCID mice (see details in Supplementary Experimental Procedures).
References
Al-Hajj M . (2007). Cancer stem cells and oncology therapeutics. Curr Opin Oncol 19: 61–64.
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF . (2003). Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA 100: 3983–3988.
Chen CZ . (2005). MicroRNAs as oncogenes and tumor suppressors. N Engl J Med 353: 1768–1771.
Cho RW, Clarke MF . (2008). Recent advances in cancer stem cells. Curr Opin Genet Dev 18: 48–53.
Croce CM, Calin GA . (2005). miRNAs, cancer, and stem cell division. Cell 122: 6–7.
Dalerba P, Cho RW, Clarke MF . (2007). Cancer stem cells: models and concepts. Annu Rev Med 58: 267–284.
Dalerba P, Clarke MF . (2007). Cancer stem cells and tumor metastasis: first steps into uncharted territory. Cell Stem Cell 1: 241–242.
Dontu G, Abdallah WM, Foley JM, Jackson KW, Clarke MF, Kawamura MJ et al. (2003). in vitro propagation and transcriptional profiling of human mammary stem/progenitor cells. Genes Dev 17: 1253–1270.
Gilmore AP . (2005). Anoikis. Cell Death Differ 12 (Suppl 2): 1473–1477.
Glinsky GV, Berezovska O, Glinskii AB . (2005). Microarray analysis identifies a death-from-cancer signature predicting therapy failure in patients with multiple types of cancer. J Clin Invest 115: 1503–1521.
Gupta PB, Chaffer CL, Weinberg RA . (2009). Cancer stem cells: mirage or reality? Nat Med 15: 1010–1012.
Ji Q, Hao X, Zhang M, Tang W, Yang M, Li L et al. (2009). MicroRNA miR-34 inhibits human pancreatic cancer tumor-initiating cells. PLoS ONE 4: e6816.
Jordan CT, Guzman ML, Noble M . (2006). Cancer stem cells. N Engl J Med 355: 1253–1261.
Mizejewski GJ . (1999). Role of integrins in cancer: survey of expression patterns. Proc Soc Exp Biol Med 222: 124–138.
Mo YY, Yu Y, Theodosiou E, Ee PL, Beck WT . (2005). A role for Ubc9 in tumorigenesis. Oncogene 24: 2677–2683.
Muller S, Hoege C, Pyrowolakis G, Jentsch S . (2001). SUMO, ubiquitin's mysterious cousin. Nat Rev Mol Cell Biol 2: 202–210.
Park SW, Hu X, Gupta P, Lin YP, Ha SG, Wei LN . (2007). SUMOylation of Tr2 orphan receptor involves Pml and fine-tunes Oct4 expression in stem cells. Nat Struct Mol Biol 14: 68–75.
Ponti D, Costa A, Zaffaroni N, Pratesi G, Petrangolini G, Coradini D et al. (2005). Isolation and in vitro propagation of tumorigenic breast cancer cells with stem/progenitor cell properties. Cancer Res 65: 5506–5511.
Pontier SM, Muller WJ . (2009). Integrins in mammary-stem-cell biology and breast-cancer progression—a role in cancer stem cells? J Cell Sci 122: 207–214.
Watanabe M, Takahashi K, Tomizawa K, Mizusawa H, Takahashi H . (2008). Developmental regulation of Ubc9 in the rat nervous system. Acta Biochim Pol 55: 681–686.
Wu F, Zhu S, Ding Y, Beck WT, Mo YY . (2009). MicroRNA-mediated regulation of Ubc9 expression in cancer cells. Clin Cancer Res 15: 1550–1557.
Yu F, Yao H, Zhu P, Zhang X, Pan Q, Gong C et al. (2007). let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell 131: 1109–1123.
Zhan M, Zhao H, Han ZC . (2004). Signalling mechanisms of anoikis. Histol Histopathol 19: 973–983.
Zhang Z, Liao B, Xu M, Jin Y . (2007). Post-translational modification of POU domain transcription factor Oct-4 by SUMO-1. FASEB J 21: 3042–3051.
Acknowledgements
This work was supported by the Natural Science Foundation of China Grant (30921140312, 30830110, 30772550, 30973396, 30525022, 30671930, 30973505), 973 (2010CB912800, 2009CB521706, 2005CB724605) Projects from the Ministry of Science and Technology of China, and Natural Science Foundation of Guangdong Province (8251008901000011).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Oncogene website
Supplementary information
Rights and permissions
About this article
Cite this article
Yu, F., Deng, H., Yao, H. et al. Mir-30 reduction maintains self-renewal and inhibits apoptosis in breast tumor-initiating cells. Oncogene 29, 4194–4204 (2010). https://doi.org/10.1038/onc.2010.167
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2010.167
Keywords
This article is cited by
-
Understanding crosstalk of organ tropism, tumor microenvironment and noncoding RNAs in breast cancer metastasis
Molecular Biology Reports (2023)
-
miR-30a targets STOX2 to increase cell proliferation and metastasis in hydatidiform moles via ERK, AKT, and P38 signaling pathways
Cancer Cell International (2022)
-
A 10-miRNA risk score-based prediction model for pathological complete response to neoadjuvant chemotherapy in hormone receptor-positive breast cancer
Science China Life Sciences (2022)
-
MicroRNA regulation of cancer stem cells in the pathogenesis of breast cancer
Cancer Cell International (2021)
-
Heterogeneity of BCSCs contributes to the metastatic organotropism of breast cancer
Journal of Experimental & Clinical Cancer Research (2021)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}