
Abstract
Understanding dryland dynamics is essential to predict future climate trajectories. However, there remains large uncertainty on the extent to which drylands are expanding or greening, the drivers of dryland vegetation shifts, the relative importance of different hydrological processes regulating ecosystem functioning, and the role of land-use changes and climate variability in shaping ecosystem productivity. We review recent advances in the study of dryland productivity and ecosystem function and examine major outstanding debates on dryland responses to environmental changes. We highlight often-neglected uncertainties in the observation and prediction of dryland productivity and elucidate the complexity of dryland dynamics. We suggest prioritizing holistic approaches to dryland management, accounting for the increasing climatic and anthropogenic pressures and the associated uncertainties.
Similar content being viewed by others


Main
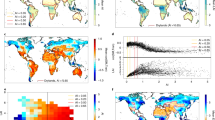
Drylands are commonly defined as regions where precipitation is substantially smaller than atmospheric water demand (as quantified by potential evapotranspiration, PET). They are the largest biome on Earth1, covering about 40% of the terrestrial land surface (Fig. 1a). Their climates are typically characterized by infrequent, seasonal and highly variable precipitation and intense solar radiation2,3. Despite their low and often discontinuous vegetation cover (Fig. 1b), drylands contribute to about 40% of global net primary productivity (NPP) (Fig. 1c) and play an important role in the global carbon budget4,5, particularly in determining the variability and long-term trend of the terrestrial CO2 sink5,6. Dryland productivity is controlled mainly by the amount and variability of precipitation7, although plant water stress can be mitigated by ecohydrological feedbacks between plant communities and hydrological processes, access to groundwater and non-rainfall water and, in cultivated areas, irrigation3,8.

a, The global distribution of drylands of different aridity classes and the percentages of the global population they support. b, Mean dryland vegetation greenness as indicated by the NDVI from moderate-resolution imaging spectroradiometer (MODIS, MOD13C2 product, 2000–2020). c, The percentage of NPP of different aridity classes compared with the global NPP (MODIS, MOD17A3 product, 2000–2020).
Drylands host more than two billion people (Fig. 1a) and provide essential ecosystem services associated with the supply of plant biomass for food, fibre and energy3,9. Therefore, vegetation productivity is also of great economic importance in the agrarian societies populating dryland regions of the world, where crop and livestock production serve as the major source of employment and livelihood. Despite their usually low ecosystem productivity, drylands are often more agriculturally productive than the wet tropics or boreal forests, which have higher ecosystem productivity. This is due to their favourable temperature and radiation conditions and, where feasible, the use of irrigation. As such, drylands have been breadbaskets for millennia, and multiple staple crops (for example, wheat) originated from drylands. In recent decades, major land-use changes occurred in global drylands, and agriculture has been expanding to meet local and global demands for food, feed and bioenergy. For example, ~225,000 km2 of tree-covered dryland were converted into other land-cover types between 1992 and 2015; 56% of that area transitioned to shrubland, while 40% was converted to cropland (Fig. 2).

a, The spatial distribution of the major land-use/land-cover types in 2015 in global drylands. Land-use/land-cover data are from European Space Agency Climate Change Initiative. b, The circos plot shows the land-use/land-cover change from 1992 to 2015. The colour of the outside circle indicates the percentage of land-cover change; the first-layer inner circle colour represents the land-use type in the corresponding year; the second-layer inner circle indicates the changed land-cover area in the unit of 1,000 km2. Coloured flows show pathways of land-use transitions from 1992 to 2015. For example, ~225,000 km2 of the tree-covered regions are converted into other land-cover types between 1992 and 2015 (orange colour in the interior circle). Among all the converted tree-cover regions, 56% of the decreased tree-cover regions (purple colour for the outside circle for the ‘Tree cover’ region in 1992) are changed into shrubland in 2015. Of all the added shrubland in 2015, 90% is from regions that were tree covered in 1992 (orange-coloured outside circle for ‘Shrub’ region in 2015), and 7% are from regions that were cropland in 1992 (red-coloured outside circle for Shrub region in 2015).
Dryland productivity trends are affected by both climate change and natural climate variability (for example, El Niño/Southern Oscillation) through changes in plant water availability driven by trends and fluctuations in both precipitation and temperature. Climate change typically induces changes in both mean state and variability of climate variables. While projected precipitation changes are still uncertain, global warming is expected to increase climate variability, including precipitation variability10,11. How drylands respond to the ongoing and future climate change will dramatically affect the trajectory and magnitude of the terrestrial carbon sink and land–atmospheric coupling. However, questions remain open on how climate change will alter the spatial extent of drylands, trends and drivers of dryland vegetation productivity and dryland contribution to the global carbon cycle.
In this Review, we provide a synthesis of the recent advances in the understanding of trends and drivers of dryland productivity and ecosystem functions. We also delve into the ongoing debates around dryland expansion, greening and vegetation response to both hydroclimatic drivers and human actions (Fig. 3). We finally highlight major knowledge gaps and suggest future research opportunities and priorities.

The complexity of dryland dynamics is elucidated in the central panel. These complexities interact with local and global climatic, ecological and land-management drivers (scheme on the left) and ultimately determine dryland productivity and functions in current and future climates (scheme on the right). The often-neglected major uncertainties in observations and predictions of dryland productivity are highlighted at the bottom.
Observed trends and debates on the future of drylands
Here we present the key observations of dryland vegetation dynamics and discuss the ongoing debates regarding dryland expansion/greening.
Observed global dryland vegetation dynamics and drivers
Spatiotemporal vegetation productivity dynamics and their drivers are of great interest to assess changes in environmental conditions in drylands. Satellite remote sensing demonstrates global dryland greening trends in the past three decades12 (Fig. 4). However, while greening was observed across the Sahel13, the Tibetan Plateau and the western United States14, large areas of the southwestern United States, southern Argentina, Kazakhstan, Mongolia, Afghanistan and regions of Australia have instead seen a decrease in vegetation cover15. Some of the observed differences in vegetation trends may be caused by different time periods selected and the sensitivity of these regions to decadal climate oscillations. Harder to detect from space are ongoing trends in plant community composition such as shrub encroachment, exogenous grass invasions16, tree die-offs17 and the increasing dominance of plants with crassulacean acid metabolism18.

a,b,d,e,g,h, The spatial distribution of vegetation productivity trends (slopes of productivity–time relationships) indicated by Global Inventory Modeling and Mapping Studies (GIMMS) LAI (LAI3g) (a), Global Land Surface Satellite (GLASS) LAI (b), GIMMS NDVI (NDVI3g) (d), VOD climate archive (VOD) (e), light-response-function-based GPP (LRF GPP) (g) and eddy covariance and light-use-efficiency-based GPP (EC-LUE GPP) (h). c,f,i, The trends of annual mean global dryland productivity anomaly (z score) based on different products: LAI3g and GLASS LAI (c), NDVI3g, VOD and MODIS NDVI (f) and LRF GPP, EC-LUE GPP and MODIS GPP (i). In c, f and i, the solid lines represent annual means and the shaded areas represent one standard deviation of the annual means. Dots in the spatial maps indicate significance at α = 0.05.
The main drivers of dryland vegetation dynamics could be broadly divided into factors related to global climate (for example, precipitation and temperature) and human actions (for example, grazing, afforestation/deforestation, agriculture, fire management, urbanization)19,20. At the global scale, drivers of greening have been related mainly to atmospheric CO2 fertilization, increased vegetation water-use efficiency (WUE) and climate warming12,21,22 (Fig. 3). While precipitation is the main driver of changes in dryland greenness globally, local controls are also important23,24,25. At the regional scale, other factors such as nitrogen deposition and land-use change also play important roles in determining vegetation dynamics20 (Fig. 3). Climate change factors such as warming, altered precipitation regimes (for example, increased variability) and increased CO2 levels can facilitate woody plant encroachment (the increasing abundance of woody plants in grasslands and savannahs) at the global scale26, while human activities such as fire suppression and overgrazing act at more local scales. Other human activities, including agriculture and deforestation, however, can locally offset the global trend of increasing woody plant abundance27. While changes in vegetation phenology as a result of climate trends and changes in plant community composition are expected to affect spatiotemporal patterns of plant growth in ecosystems, their impact on the productivity of the global drylands remains unclear. Modelling studies constrained by field observations28 found an improvement in the model performance of grassland productivity simulation when accounting for trends in semi-arid grassland phenology29. These results suggest that a future shift towards both earlier growing season onset and delayed senescence could compensate for drought-induced reductions in summer grass cover and productivity, resulting in widespread increases in grassland fractional cover over the coming century across the majority of North American grasslands28.
Dryland expansion debates under future climates
It has been argued that climate change will increase the aridity of existing drylands and/or lead to their expansion, with negative impacts on ecosystem productivity and livelihoods30,31. These trends, however, are still debated32. According to the dryland expansion hypothesis, the expected intensification of dryland aridity will lead to vegetation loss and reduced primary productivity. At the same time, there is empirical evidence of dryland greening, suggesting an increase in productivity consistent with the known effects of increased atmospheric CO2(refs.21,33). Despite the positive CO2 effect on WUE, drylands are expected to expand in a warming climate30,31 according to model estimates of the aridity index (AI = precipitation/PET). Indeed, model projections show that warming is expected to induce a global increase in PET (~5% °C−1) outpacing the global increase in precipitation on land (1–2% °C−1). Moreover, in the specific case of drylands (regions with AI < 0.65), precipitation has been observed to decrease in the past 40 years, with a few exceptions34. Thus, under climate warming scenarios, AI is projected to decrease worldwide, particularly in drylands, which are also predicted to expand.
Such conclusions, however, have been recently challenged because AI does not seem to be suitable for the prediction of dryland extent and aridity under future climate scenarios. In fact, the calculation of AI using the Penman–Monteith PET equation does not consider vegetation response to higher CO2 concentration. If this response is dominated by stomatal closure, neglecting CO2 effects may lead to an overestimate of PET response, aridity increase and dryland expansion under climate warming35. Lastly, changes in soil dryness are ultimately driven by the soil water balance through differences between precipitation and actual evapotranspiration (ET) instead of PET. ET’s complementary relationship (the decrease in ET with increasing values of PET) has been invoked to argue that warming is not necessarily associated with drying trends, consistent with palaeoclimatic evidence36. New aridity metrics developed to address the shortcomings of AI suggest that climate warming is not expected to substantially change the spatial extent of drylands35. Interestingly, a recent study found that over the past 30 years, vegetation growth in the Northern Hemisphere showed increasing water constraints37, suggesting that even without invoking dryland expansion under climate warming, vegetation growth in both drylands and nearby areas may suffer from increasing water limitations.
Complexity in the water–productivity relationship in drylands
Water availability and its timing play a pivotal role in dryland dynamics and productivity. Dryland response to climate change has typically been investigated with reference to trends in mean climate variables (for example, rainfall and soil moisture), while the effect of their variability (for example, seasonality, interannual variability) received much less attention24,26,38. Even less is known about the role of variability in non-rainfall water inputs (mainly fog and dew) and groundwater fluctuations39,40 on dryland vegetation dynamics. In this section, we highlight these less studied but equally important factors affecting dryland productivity globally and at a regional scale.
Interannual rainfall variability and productivity
Dryland climates are characterized by a strong intra- and interannual variability of precipitation, associated with precipitation intermittency, seasonality and year-to-year fluctuations. In arid and semi-arid regions, precipitation typically occurs clustered in one (or more) rainy season(s) interrupted by dry periods with low or no rain. Rainy seasons exhibit a few scattered events separated by rainless days. Interannual variability relative to average totals, which is often expressed by the coefficient of variation of annual precipitation, is particularly strong in drylands and tends to increase with aridity, as shown by trends in the coefficient of variation along precipitation gradients41. Most of the interannual variability results from changes in the number of rainy days rather than from changes in the average precipitation amount on rainy days39.
Overall, interannual precipitation variability decreases above-ground NPP and the terrestrial carbon sink in dryland ecosystems with mean annual precipitation >300 mm, while aboveground NPP increases with precipitation variability in drier climates as vegetation benefits from the wet anomalies38,40. Within this general pattern, an increase in interannual precipitation variance typically increases the interannual variability of vegetation cover or plant productivity. The effects of random interannual precipitation fluctuations could be less ‘trivial’ than this when they interact with nonlinearities in ecosystem dynamics and induce newly organized states, bifurcations and spatiotemporal patterns that would not exist in the absence of environmental variability42. For example, an increase in precipitation variability may lead to the emergence of alternative stable states in soil moisture dynamics, indicating that a certain region may have a higher likelihood to be in a dry or in a relatively wet state while intermediate conditions have a lower probability of occurrence. The opposite can also occur. Interannual precipitation variability may remove bistability in systems that would otherwise exhibit two alternative stable configurations and stabilize vegetation dynamics in an intermediate state42. Coupled vegetation–climate models have shown how, in the absence of interannual climate variability, the savannah belts across the Sahel and southern Africa tend to exhibit bistable dynamics with two preferential states of ‘desert’ and ‘vegetated’ land. This bistable behaviour, however, can be inhibited by interannual climate fluctuations43, which stabilize the system in a state with intermediate vegetation density (and productivity), thereby enhancing its resilience41.
In dryland ecosystems, interannual precipitation variability can also induce vegetation patterns, including the periodic spotted, banded and labyrinthine vegetation configurations widely documented in drylands44. A major implication of pattern formation is its ability to enhance the resilience and productivity of plant ecosystems45. Past research, however, has strongly relied on model simulations with only few manipulative experiments46. Because self-organized patterns with similar geometries can result from models that account for different processes, the mechanisms underlying pattern formation and the role of interannual precipitation variability still need to be conclusively assessed44.
Precipitation variability plays an important role also in the dynamics of dryland plant communities and has been invoked to explain changes in woody plant dominance47. Interannual precipitation fluctuations are also expected to have a stronger impact on annual than perennial species, thereby potentially reshaping the composition of grass communities48.
The effect of intra-annual rainfall variability on productivity and pulses
The intra-annual variability of precipitation is also an important determinant of vegetation composition and productivity. Tree cover increases with increasing frequency of rainy days and decreasing precipitation intensity49, and similarly, productivity increases when the precipitation events occur regularly, thus limiting the effects of water stress. In some drylands, even though average precipitation amount does not change, there are alterations in seasonal, daily or sub-daily rainfall regime characteristics (frequency, duration, intensity)50,51. These changes could have profound impacts on vegetation functioning and species composition. For example, C3 plant abundance increases under low rainfall and high temperatures, despite C4 plants being better adapted to such conditions52. This surprising finding can be explained by higher precipitation during cooler months—when C3 grasses are most active—during extreme drought years52. Precipitation variability may also influence ecosystems through interactions with the temperature regime. In addition, the physiological impact of rising CO2 (through higher WUE) may act by downregulating or elevating the sensitivity of dryland plants to precipitation variability53.
Intra-annual precipitation variability also triggers biogeochemical pulses. In drylands, prolonged dry periods between rain events or seasonal droughts are followed by marked increases in soil moisture at rewetting that re-establish microbial catabolic activity and plant gas exchanges (over hours to days), followed by a growth pulse in both microbial communities (hours to days) and plants (days to weeks)54,55. As microbial activity resumes, a pulse of respiration and nutrient release ensues. These pulses are important because they are responsible for a large fraction of the annual GHG exchanges (thus affecting soil carbon stocks) and for supplying nutrients fuelling plant productivity56. While the occurrence of these pulses is well known, there remain several open questions. What drives the pulse in microbial activity? Are the pulses of respiration, microbial growth and nutrient release synchronous, indicating efficient resource use and nutrient retention in the ecosystem? How are pulse dynamics changing with the ongoing increasing precipitation variability and lengthening of dry periods?
Heterotrophic respiration pulses are caused by several simultaneous processes, ranging from mineralization of dissolved organic compounds accumulated during the dry period to consumption of microbial by-products or necromass to physical disruption of aggregates that were protecting organic matter during the dry period57. These processes are intensified by large changes in moisture after a long dry period, leading to the hypothesis that ongoing increasing precipitation variability is increasing the contribution of respiration pulses in drylands58. Importantly, microbial growth at rewetting might be de-coupled from respiration as microbial growth recovery often lags behind respiration after long dry periods59. This could lead to larger carbon and nutrient losses as dry periods lengthen because carbon and nutrients are not efficiently retained in the microbial biomass. However, microbial communities can adapt to increasing precipitation variability, thereby tightening carbon and nutrient cycles59.
As microbes are reactivated at rewetting, nitrogen and phosphorus mineralization rates increase60. The released inorganic nutrients can be leached, volatilized or taken up by plants. If mineralization is faster than uptake, nutrients can accumulate (for example, nitrate) and be lost61. It is therefore critical for the retention of nutrients that microbial and plant activity are synchronized, but that is often not the case, especially after a dry season or long dry period when plant recovery is much slower than microbial recovery55. Moreover, with increasing aridity, geochemical and biological processes are likely to change at different rates, promoting, for example, phosphorus accumulation due to continued weathering when plant phosphorus uptake is low, and nitrogen depletion as organic matter production and mineralization are both inhibited under dry conditions62. Therefore, ongoing precipitation intensification and lengthening of dry periods can decouple nutrient availability and utilization, as well as create nutrient imbalances, thereby promoting nutrient losses and negatively affecting dryland productivity.
Fog and dew impacts on vegetation and ecosystem functions
With water being the predominant limiting resource in drylands, the form, rate, and timing of water input are crucial to how ecosystems can utilize and respond to water availability63. While the role of rainfall is undisputed, in many dryland regions, small but critical amounts of fog and dew are also essential for ecosystem productivity and function64,65. Fog and dew are the least studied components of the hydrological cycle in drylands8. Overlooking these non-rainfall components could lead to inaccurate results. For example, climate warming experiments commonly use infrared heater warming systems to simulate warming conditions, which can greatly reduce dew formation66.
Although the input of dew and fog is rarely a limiting factor for species or biome distributions, it can have a measurable impact on carbon and water fluxes by increasing the leaf water potential and impacting the temperature, albedo and local vapour pressure deficit (VPD) of the canopy, and providing a latent heat ‘sink’67. Fog and dew allow plants to retain more moisture in cells, and soil moisture remains higher, increasing plant resilience during hot and dry conditions65. There is also a long history of human utilization of fog and dew to meet societal needs68. Fog harvesting systems in the Atacama desert of Chile and Peru have been used for water supply and crop irrigation69, and there are reports of natural dew collection in other desert ecosystems such as the Negev of Israel70.
Despite the importance of fog and dew to the functioning of different dryland ecosystems (Supplementary Fig. 1), there are important gaps in knowledge regarding the magnitude of fog and dew formation under different climatic conditions and the extent that fog and dew impact the water, carbon and energy fluxes at the canopy scale. Research has focused largely on a limited number of field measurements at the leaf or plant level. Lack of knowledge of the canopy-scale effects of fog and dew makes it difficult to constrain the degree of fog and dew contribution to ecosystem functioning and evaluate the effects of climate change on water limitations. In fact, rising night-time and daytime VPD with climate warming71 is expected to generate a global-scale decline in fog and dew occurrence and duration66,72, possibly contributing to the increasing water constraints on vegetation growth across diverse biomes observed over the past 30 years37.
Dryland bistability and desertification
The response of dryland ecosystems to changes in climate and land use may be nonlinear and undergo relatively abrupt and often irreversible transitions to a different configuration, suggesting that the underlying dynamics might have two (alternative) stable states. Such a bistable behaviour is typically attributed to positive feedbacks between vegetation and the physical environment73. For example, plants can modify the surrounding environment, creating their own habitat. This phenomenon is widespread in drylands where plants can reduce soil erosion, enhance infiltration, enhance fog and nutrient deposition or prevent soil salinization, thereby favouring their own survival and growth73,74. As a result of these feedbacks, dryland ecosystems have a limited resilience: if disturbed beyond a critical threshold, they can shift to an undesirable state characterized by land degradation and loss of ecosystem services or productivity39.
The term desertification is often used to denote the critical transition of dryland ecosystems to undesirable ‘desert-like’ conditions75,76. This notion suffers from inherent ambiguity resulting from the fact that desert landscapes can be very different, and the shift to desert-like conditions can result from a variety of drivers and processes. Desertification may consist of a loss of soil productivity due to erosion, salinization or soil toxicity. It may also result from aridification trends sustained by vegetation loss or from shifts in plant community composition associated with woody plant encroachment or biological invasions75. However, some of these undesired shrubland states may have higher productivity and biodiversity than their grassland counterpart77. Thus, ‘desertification’ and ‘land degradation’ often refer to a loss of ecosystem services and economic productivity and not necessarily a shift to ecologically unproductive states75. Overall, there is some ambiguity in what desertification actually entails, which is contributed partly by the inconsistent definitions of this phenomenon75,78.
Regional differences in dryland productivity changes
Depending on the major geographic and hydroclimatic factors determining dryland occurrence, drylands may respond differently to climate change. Regions with divergent air masses and subsiding air such as the subtropics (for example, the Sahel, the Arabian Peninsula, the Kalahari, Central Australia and the Chihuahuan and Sonoran deserts) are expected to become drier under climate warming, a phenomenon that has been ascribed to (1) the enhancement of existing subtropical aridity zones and/or (2) the poleward expansion of the subtropics, a phenomenon that is stronger in the Southern Hemisphere79. Other drylands such as those in central Asia are located far from ocean sources of atmospheric moisture (the so-called continentality effect). Because local transpiration can be an important contributor to atmospheric moisture in these drylands, their precipitation regime can be altered by local vegetation cover and land-use changes. Coastal deserts (for example, the Namib and the Atacama) exhibit frequent water inputs through fog and dew. In such systems, changes in fog and dew regimes are expected to drive changes in ecosystem productivity. Specifically, the amount of fog and dew water input is expected to decrease as a result of climate warming and its effects on condensation. The productivity of some drylands is co-limited by water and temperature (for example, the Qinghai–Tibetan Plateau, the Gobi Desert and the Chihuahuan Desert). As such, productivity in these regions could benefit from warming (at least in the short term). For other dryland regions where snowmelt from high mountain chains is the primary source of recharge (for example, western South America and western North America), warming leads to an increase in the fraction of precipitation falling as rain compared with snow, resulting in a decline in spring and summer streamflow, groundwater recharge and seasonal water storage in these regions80. Dryland soil and land management may also play a role in determining regional differences and divergent responses of drylands to climate trends (more details are in Supplementary Note 1).
Dryland agriculture and management
Some drylands are considered ‘marginal lands’ because they exhibit relatively low ecosystem productivity or biodiversity81. However, agricultural and natural ecosystems differ in productivity. Agriculture in drylands can be productive, particularly in the presence of irrigation and fertilization. Drylands sustain livelihoods and wildlife and provide a sense of place to rural communities, including indigenous peoples that have stewarded these lands often under communal or traditional uses for generations. Dryland agriculture supplies much of the world’s food and fibres, providing about 44% of the global agricultural land and 60% of global food production82. Nevertheless, the dependence of primary productivity on water resources may limit crop production in drylands, where reliable and sufficient water availability for irrigation is necessary to stave off the effects of water and heat stress and achieve high and stable crop yields. Thus, yield gaps are often high in drylands, particularly in rainfed agriculture83.
Cropland area in drylands has increased by about 10% from 2003 to 2019 (Fig. 5) as a result of both local needs and global markets through international trade84. This global telecoupling between crop demand and production regions is associated with a global displacement of land use85. In the aftermath of the 2008 food and financial crisis, large tracts of land worldwide have been targeted by large-scale land acquisitions (LSLAs), allegedly with the aim of increasing food and energy production or curbing GHG emissions, although often for mere financial speculation86. In the past few decades, cropland and rangeland in North and sub-Saharan Africa, South America and central and southeast Asia have expanded at the expense of natural ecosystems (Fig. 5), to some extent as a result of LSLAs87,88. In the mid-latitude drylands of Russia, Ukraine and Kazakhstan, land investors have restored abandoned croplands to their previous agricultural use. LSLAs are also contributing to a transition from small-scale/subsistence farming to large-scale commercial agriculture in dryland regions of the developing world. These transitions may have negative socioenvironmental impacts on rural livelihoods, land stewardship, common property regimes, natural capital, water resources and soil conservation89.

Dryland agriculture is expected to be negatively affected by climate change, with a decline in both crop and livestock productivity as a result of temperature extremes (hot and cold), decreasing precipitation, lowering groundwater tables and increasing land degradation78. By 2050, under the ‘middle of the road’ shared socioeconomic pathway (SSP2) at 1.5 °C warming, 178 million people are projected to be vulnerable to water stress, drought intensification and habitat degradation in dryland regions78. Climate change will also reshape the global distribution of regions suitable for sustainable expansion of irrigation, while requiring increased annual water-storage capacity90. Despite these challenges, drylands are expected to remain crucial for global food and bioenergy production, with further expansion of agriculture and intensification of existing crop production, mostly in the tropics78. Agricultural intensification in drylands will require investments in precipitation conservation and irrigation91 to improve crop productivity in regions with big yield gaps. Nevertheless, a sustainable expansion of irrigation can be achieved only in semi-arid to sub-humid dryland regions, while in drier areas, water resources will be sufficient to irrigate only part of the land or meet part of the crop water demand (that is, deficit irrigation will be needed). Major increases in cropland productivity via irrigation expansion are thus expected in sub-Saharan Africa and Southern Eurasia90 unless inadequate land and water governance prevents investments in irrigation.
In addition to climate change, evolving dietary habits are placing further pressure on cropland and rangeland. Meat consumption in developing countries is expected to increase by 125% from 2005–2007 to 205092. The growth in livestock production observed in the past few decades is projected to continue in the near future and expand to dryland regions of the developing world, for example, as now occurring in the Asian dryland belt93. Grazing systems are already threatened by combinations of land degradation, declining grassland productivity and overgrazing94, possibly leading to regime shifts to low-productivity states particularly in drylands. Nevertheless, smallholders operating mixed crop–livestock systems in developing countries are projected to remain the main producers of ruminant livestock by 205095.
Uncertainties and opportunities
Here we present the key uncertainties in dryland productivity observation, modelling and driver attribution. We also highlight the future opportunities.
Uncertainties in observing dryland productivity
Despite recent advances in the study of dryland vegetation, some large uncertainties remain in assessing its productivity. One of the major sources of uncertainty comes from the choice of dryland productivity indicator. For example, while vegetation indices (for example, normalized difference vegetation index, NDVI) are sensitive to chlorophyll concentrations and canopy cover fraction, their effectiveness is limited in areas with low vegetation cover and large soil background96. Analyses of global vegetation products showed both greening of dryland vegetation and a hidden global browning trend, depending on underlining canopy density97. There are also uncertainties arising from climate variability and the sensitivity of temporal trend analysis to starting and ending periods25,98. Finally, uncertainties in satellite imagery interpretations and lack of high-quality ground observations cause misclassification of land-cover types that sustains ongoing debates on changes in dryland vegetation cover99,100.
Uncertainties in predicting dryland productivity
The role drylands play in the global carbon cycle is often evaluated using process-based global terrestrial biosphere models (TBMs), but uncertainties in TBM predictions remain large101,102,103. For example, the recent model intercomparison TRENDY v7, based on a suite of 14 TBMs, showed that all models underestimated both mean annual net ecosystem exchange and its interannual variability when compared with in situ CO2 flux measurements in the southwestern United States102. This net ecosystem exchange underestimate was caused by the models showing a too weak response of vegetation growth and gross primary productivity (GPP) to changes in plant water availability. Moreover, in both Australia and the southwestern United States, models dramatically underestimated interannual variability in GPP (Fig. 6). The uncertainties in the magnitude of the interannual variability of Australian net biome production (NBP) simulated by the TRENDY v8 models for 1901–2018 led to an accumulated ~14 PgC spread in NBP across models, and no agreement on whether the ecosystem is a net carbon sink or source101.

A comparison of annual GPP and WUE from a suite of global TBMs from the TRENDY v.7 model intercomparison project against eddy covariance data from 19 flux tower sites in Australia (orange symbols and bars) and the southwestern United States (SW US, blue symbols and bars). a, Site locations. b, Slope of the linear regression between annual GPP simulated by all 14 TRENDY v.7 models and observed annual GPP across all Australian sites. c, Same as b but for SW US sites. A slope of 0.0 is shown in dark grey dashed lines and a slope of 1.0 is shown in black dashed lines. Individual sites are denoted by circle markers. For all boxplots, the boxes show the interquartile range and median of the spread across all sites, and the whiskers extend to 1.5 times the interquartile range (~99% of the distribution). Outliers outside this range are shown with black crosses. The slopes in b and c are close to zero, indicating the interannual variability was not well captured by the models. d, Slope of the relationship between monthly GPP (g m−2 month−1 C) and monthly ET (mm month−1) (a measure of ecosystem WUE) across all months and all Australian sites for 12 of the TRENDY v.7 models (orange bars) compared with the observed ecosystem WUE (grey bars). e, Same as d but for SW US sites. Light grey dashed lines in d and e show the median value of the observed ecosystem WUE. Australian sites include AU-ASM, AU-Cpr, AU-DaS, AU-Dry, AU-Gin, AU-How and AU-Stp, and the GPP and ET data were taken from FLUXNET 2015 database122. SW US sites include US-Fuf, US-Mpj, US-Wjs, US-Vcm, US-Vcp, US-Ses, US-Seg, US-Aud, US-SRM, US-SRG, US-Wkg and US-Whs, and the GPP and ET data were obtained from the site principal investigators (see ref. 102 for further details on data processing; for the full list of site names, see Supplementary Table 1). TRENDY TBMs include CLM v.5.0, JULES, ORCHIDEE v.2.0, ORCHIDEE-CNP, OCN, JSBACH, CABLE-POP, ISAM, CLASS-CTEM, SDGVM, LPX, LPJ, DLEM and SURFEX. TRENDY v.7 S3 simulations were used, including changing climate forcing, rising atmospheric CO2 concentrations and land-use change (see ref. 102 for further details).
Missing or misrepresented processes in TBMs cause inaccurate long-term carbon uptake and accumulation estimates. Incorrect vegetation sensitivity to changes in water availability undermine predictions of productivity interannual variability101,102,103,104. Uncertain satellite-derived estimates of plant functional type fractional cover lead to large intermodel spread in dryland productivity and water fluxes105. Discrepancies exist also between observed and modelled fractional cover. TBMs generally do capture well the daily to interannual variability of dryland in situ soil moisture and ET (as a proxy of plant water availability)103,106, but model–data discrepancies remain when considering the partitioning of ET at dryland sites into its constituent transpiration and bare soil evaporation components106. Future TBM evaluation studies should test processes related to dryland vegetation composition, structure and functioning, particularly in relation to plant water uptake and use and fire dynamics. More elaborated discussion on this topic can be found in Supplementary Note 2.
Uncertainties in understanding dryland productivity drivers
Despite consistency in broad trends in dryland vegetation changes, large uncertainties remain in explicitly attributing the drivers of observed dryland vegetation dynamics107, in part due to the uncertainties in the TBMs used in attribution studies. A number of open questions and sources of uncertainties in model estimates remain. How is dryland productivity affected by climate oscillations such as El Niño/Southern Oscillation107? To what extent can rising CO2 ameliorate plant water stress in water-scarce conditions108, considering that CO2 fertilization enhances plant photosynthesis and leaf area index (LAI)109 and transpiring biomass110, while also reducing stomatal conductance and thus transpiration per unit leaf area21,111? Do land–atmosphere feedbacks increase the concurrence of high VPD and low soil moisture, consequently increasing plant water stress and limiting plant growth112,113? In addition, model-based dryland productivity driver analyses do not always agree with remote-sensing-based studies15,107,109, probably because of the lack of integration of high-resolution data with field observations107. Understanding the key drivers of dryland vegetation dynamics is urgently needed in the light of the projected increasing drought frequency and severity114.
There are also notable biases of prediction models in simulating extreme precipitation115 and drizzle116, with only small improvements of Coupled Model Intercomparison Project phase 6 over phase 5117 in precipitation modelling. The uncertainty of predicted precipitation also enhances the uncertainty of dryland productivity estimates and predictions into the future. In addition, groundwater trends and fluctuations due to climate change and human actions are not well constrained in many drylands118. This has large implications on the understanding and projection of dryland functioning because access to subsurface water, especially groundwater, contributes to dryland productivity119.
Future opportunities and research priorities
While uncertainties remain, there have been important recent advances in observation technologies, modelling approaches and statistical tools to quantify vegetation productivity and attribute it more accurately to different drivers. The development of ground observation networks and remote-sensing technology focusing on both land cover/land use and vegetation productivity provide unprecedented opportunities for reconciling the differences in trends of dryland vegetation dynamics across spatial scales. The increasingly available ground observations through meteorological and flux tower networks reduce uncertainties from satellite remote-sensing retrievals. At the same time, novel satellite-based data allow mapping vegetation structure—not only vegetation greenness (Supplementary Note 3). TBM testing, optimization and developments are under way. Specific processes that require TBM advances include vegetation sensitivity to changes in water availability, a more dryland-specific phenology scheme, disturbance representation and the relative control of VPD and soil moisture on dryland vegetation productivity. Developments in remote-sensing approaches and deployment of networks of in situ dryland observations will be crucial in this regard. These steps will increase the reliability of TBMs to predict dryland productivity and the role of drylands in the global carbon cycle under changing climate.
Besides scientific research, more holistic dryland management and planning is required to avoid land degradation. It is imperative that we address these research and management challenges urgently because drylands are an important player in the Earth system, provide essential ecosystem services to human society and are undergoing dramatic, and potentially irreversible, changes due to climate trends and land use. New observational platforms and rapidly improving process-based models that offer novel insights on the complexity of dryland dynamics can help reduce uncertainties and suggest ways to overcome these challenges.
Data availability
The advanced very high-resolution radiometer GIMMS-NDVI3g is available at https://ecocast.arc.nasa.gov/data/pub/gimms/3g.v0. Global Land Surface Satellite (GLASS) LAI can be obtained from http://www.glass.umd.edu/Download.html. The aridity index dataset is available at https://cgiarcsi.community/data/global-aridity-and-pet-database/. Moderate-resolution imaging spectroradiometer (MODIS) based EVI and GPP datasets are available from the NASA Land Processes Distributed Active Archive Center at https://lpdaac.usgs.gov. The MODIS NPP dataset is available from https://lpdaac.usgs.gov/products/mod17a3hgfv006. Ku-band VOD datasets are available from https://zenodo.org/record/2575599#.XyLqfLdME0M. European Space Agency- (ESA-) based land-use/land-cover product is available from https://www.esa-landcover-cci.org/. Light response function- (LRF-) based GPP data are available from https://doi.org/10.17894/ucph.b2d7ebfb-c69c-4c97-bee7-562edde5ce66. Light-use efficiency model- (EC-LUE-) based GPP data can be obtained from https://doi.org/10.6084/m9.figshare.8942336.v3. Eddy covariance flux tower data are available for SW US sites from the AmeriFlux database (http://ameriflux.lbl.gov) and for Australian sites from the FLUXNET 2015 database (https://fluxnet.org/data/fluxnet2015-dataset/). More information on the TRENDY MIP and related simulations is available at https://sites.exeter.acuk/trendy/.
References
Schimel, D. S. Drylands in the Earth system. Science 327, 418–419 (2010).
Whitford, W. G. Ecology of Desert Systems (Academic Press, 2002).
D’Odorico, P., Porporato, A. & Runyan, C. W. Dryland Ecohydrology Vol. 9 (Springer, 2019). A comprehensive introduction to dryland ecohydrology.
Lal, R. Carbon cycling in global drylands. Curr. Clim. Change Rep. 5, 221–232 (2019).
Ahlström, A. et al. The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science 348, 895–899 (2015). Illustrates the role drylands play in determining the variability and long-term trend of the terrestrial CO2 sink.
Poulter, B. et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle. Nature 509, 600–603 (2014). Illustrates the role drylands play in determining the variability of the terrestrial CO2 sink.
Maestre, F. T. et al. Structure and functioning of dryland ecosystems in a changing world. Annu. Rev. Ecol. Evol. Syst. 47, 215–237 (2016). A comprehensive review of dryland structure and functioning.
Wang, L., Kaseke, K. F. & Seely, M. K. Effects of non-rainfall water inputs on ecosystem functions. WIREs Water 4, e1179 (2017). Highlights the often-ignored role of non-rainfall water inputs to dryland ecosystem dynamics.
Li, C. et al. Drivers and impacts of changes in China’s drylands. Nat. Rev. Earth Environ. 2, 858–873 (2021).
Thornton, P. K., Ericksen, P. J., Herrero, M. & Challinor, A. J. Climate variability and vulnerability to climate change: a review. Glob. Change Biol. 20, 3313–3328 (2014).
IPCC Climate Change 2022: Impacts, Adaptation, and Vulnerability (eds Pörtner, H.-O. et al.) (Cambridge Univ. Press, 2022).
Gonsamo, A. et al. Greening drylands despite warming consistent with carbon dioxide fertilization effect. Glob. Change Biol. 27, 3336–3349 (2021).
Kaptué, A. T., Prihodko, L. & Hanan, N. P. On regreening and degradation in Sahelian watersheds. Proc. Natl Acad. Sci. USA 112, 12133–12138 (2015).
Brookshire, E. J., Stoy, P. C., Currey, B. & Finney, B. The greening of the Northern Great Plains and its biogeochemical precursors. Glob. Change Biol. 26, 5404–5413 (2020).
Song, X.-P. et al. Global land change from 1982 to 2016. Nature 560, 639–643 (2018).
Ravi, S. et al. Biological invasions and climate change amplify each other’s effects on dryland degradation. Glob. Change Biol. 28, 285–295 (2022).
Allen, C. D., Breshears, D. D. & McDowell, N. G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere https://doi.org/10.1890/ES15-00203.1 (2015).
Yu, K. et al. The competitive advantage of a constitutive CAM species over a C4 grass species under drought and CO2 enrichment. Ecosphere 10, e02721 (2019).
Fensholt, R. et al. in Remote Sensing Time Series (eds Kuenzer, C. et al.) 183–292 (Springer, 2015).
Andela, N., Liu, Y., Van Dijk, A., De Jeu, R. & McVicar, T. Global changes in dryland vegetation dynamics (1988-2008) assessed by satellite remote sensing: comparing a new passive microwave vegetation density record with reflective greenness data. Biogeosciences 10, 6657–6676 (2013).
Lu, X., Wang, L. & McCabe, M. F. Elevated CO2 as a driver of global dryland greening. Sci. Rep. 6, 20716 (2016).
Venter, Z., Cramer, M. & Hawkins, H.-J. Drivers of woody plant encroachment over Africa. Nat. Commun. 9, 2272 (2018).
Ukkola, A. M. et al. Annual precipitation explains variability in dryland vegetation greenness globally but not locally. Glob. Change Biol. 27, 4367–4380 (2021).
Zhang, W., Brandt, M., Tong, X., Tian, Q. & Fensholt, R. Impacts of the seasonal distribution of rainfall on vegetation productivity across the Sahel. Biogeosciences 15, 319–330 (2018).
Fensholt, R. & Rasmussen, K. Analysis of trends in the Sahelian ‘rain-use efficiency’ using GIMMS NDVI, RFE and GPCP rainfall data. Remote Sens. Environ. 115, 438–451 (2011).
Zhang, W. et al. Ecosystem structural changes controlled by altered rainfall climatology in tropical savannas. Nat. Commun. 10, 671 (2019).
Brandt, M. et al. Reduction of tree cover in West African woodlands and promotion in semi-arid farmlands. Nat. Geosci. 11, 328–333 (2018).
Hufkens, K. et al. Productivity of North American grasslands is increased under future climate scenarios despite rising aridity. Nat. Clim. Change 6, 710–714 (2016).
Choler, P., Sea, W., Briggs, P., Raupach, M. & Leuning, R. A simple ecohydrological model captures essentials of seasonal leaf dynamics in semi-arid tropical grasslands. Biogeosciences 7, 907–920 (2010).
Huang, J., Yu, H., Dai, A., Wei, Y. & Kang, L. Drylands face potential threat under 2 °C global warming target. Nat. Clim. Change 7, 417–422 (2017).
Huang, J., Yu, H., Guan, X., Wang, G. & Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 6, 166–171 (2016).
Lian, X. et al. Multifaceted characteristics of dryland aridity changes in a warming world. Nat. Rev. Earth Environ. 2, 232–250 (2021). Provides a comprehensive analysis on the dryland expansion debates.
Fatichi, S. et al. Partitioning direct and indirect effects reveals the response of water-limited ecosystems to elevated CO2. Proc. Natl Acad. Sci. USA 113, 12757–12762 (2016).
Daramola, M. T. & Xu, M. Recent changes in global dryland temperature and precipitation. Int. J. Climatol. 42, 1267–1282 (2022).
Berg, A. & McColl, K. A. No projected global drylands expansion under greenhouse warming. Nat. Clim. Change 11, 331–337 (2021).
Berg, A. & Sheffield, J. Climate change and drought: the soil moisture perspective. Curr. Clim. Change Rep. 4, 180–191 (2018).
Jiao, W. et al. Observed increasing water constraint on vegetation growth over the last three decades. Nat. Commun. 12, 3777 (2021). This study found that vegetation growth in the Northern Hemisphere is becoming increasingly water limited.
Gherardi, L. A. & Sala, O. E. Effect of interannual precipitation variability on dryland productivity: a global synthesis. Glob. Change Biol. 25, 269–276 (2019).
D’Odorico, P. & Bhattachan, A. Hydrologic variability in dryland regions: impacts on ecosystem dynamics and food security. Phil. Trans. R. Soc. B 367, 3145–3157 (2012).
Hou, E. et al. Divergent responses of primary production to increasing precipitation variability in global drylands. Glob. Change Biol. 27, 5225–5237 (2021).
Ritter, F., Berkelhammer, M. & Garcia-Eidell, C. Distinct response of gross primary productivity in five terrestrial biomes to precipitation variability. Commun. Earth Environ. 1, 34 (2020).
Ridolfi, L., D’Odorico, P. & Laio, F. Noise-Induced Phenomena in the Environmental Sciences (Cambridge Univ. Press, 2011).
Zeng, N. & Neelin, J. D. The role of vegetation–climate interaction and interannual variability in shaping the African savanna. J. Clim. 13, 2665–2670 (2000).
Borgogno, F., D’Odorico, P., Laio, F. & Ridolfi, L. Mathematical models of vegetation pattern formation in ecohydrology. Rev. Geophysics 47, RG1005 (2009).
van de Koppel, J. & Rietkerk, M. Spatial interactions and resilience in arid ecosystems. Am. Nat. 163, 113–121 (2004).
Lefever, R. & Lejeune, O. On the origin of tiger bush. Bull. Math. Biol. 59, 263–294 (1997).
Gherardi, L. A. & Sala, O. E. Enhanced precipitation variability decreases grass- and increases shrub-productivity. Proc. Natl Acad. Sci. USA 112, 12735–12740 (2015). Highlights the role of precipitation varibility in plant community composition in drylands.
Cleland, E. E. et al. Sensitivity of grassland plant community composition to spatial vs. temporal variation in precipitation. Ecology 94, 1687–1696 (2013).
Good, S. P. & Caylor, K. K. Climatological determinants of woody cover in Africa. Proc. Natl Acad. Sci. USA 108, 4902–4907 (2011).
Lu, X., Wang, L., Pan, M., Kaseke, K. F. & Li, B. A multi-scale analysis of Namibian rainfall over the recent decade—comparing TMPA satellite estimates and ground observations. J. Hydrol. Reg. Stud. 8, 59–68 (2016).
Franz, T., Caylor, K., Nordbotten, J., Rodriguez-Itubre, I. & Celia, M. An ecohydrological approach to predicting regional woody species distribution patterns in dryland ecosystems. Adv. Water Res. 33, 215–230 (2010).
Knapp, A. K., Chen, A., Griffin-Nolan, R. J., Baur, L. E. & Smith, M. Resolving the Dust Bowl paradox of grassland responses to extreme drought. Proc. Natl Acad. Sci. USA 117, 201922030 (2020).
Ukkola, A. M. et al. Reduced streamflow in water-stressed climates consistent with CO2 effects on vegetation. Nat. Clim. Change 6, 75–78 (2016).
Austin, A. T. et al. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141, 221–235 (2004). Illustrates the close linkage between water pulses and biogeochemical cycles in drylands.
Schwinning, S. & Sala, O. E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 141, 211–220 (2004).
Collins, S. L. et al. A multiscale, hierarchical model of pulse dynamics in arid-land ecosystems. Annu. Rev. Ecol. Evol. Syst. 45, 397–419 (2014).
Barnard, R. L., Blazewicz, S. J. & Firestone, M. K. Rewetting of soil: revisiting the origin of soil CO2 emissions. Soil Biol. Biochem. 147, 107819 (2020).
Manzoni, S. et al. Rainfall intensification increases the contribution of rewetting pulses to soil heterotrophic respiration. Biogeosciences 17, 4007–4023 (2020).
Leizeaga, A., Meisner, A., Rousk, J. & Bååth, E. Repeated drying and rewetting cycles accelerate bacterial growth recovery after rewetting. Biol. Fertil. Soils 58, 365–374 (2022).
Gao, D. et al. Responses of soil nitrogen and phosphorus cycling to drying and rewetting cycles: a meta-analysis. Soil Biol. Biochem. 148, 107896 (2020).
Homyak, P. M., Allison, S. D., Huxman, T. E., Goulden, M. L. & Treseder, K. K. Effects of drought manipulation on soil nitrogen cycling: a meta-analysis. J. Geophys. Res. Biogeosci. 122, 3260–3272 (2017).
Delgado-Baquerizo, M. et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 502, 672–676 (2013).
Nippert, J. B., Knapp, A. K. & Briggs, J. M. Intra-annual rainfall variability and grassland productivity: can the past predict the future? Plant Ecol. 184, 65–74 (2006).
Kaseke, K. F., Wang, L. & Seely, M. K. Nonrainfall water origins and formation mechanisms. Sci. Adv. 3, e1603131 (2017).
Dawson, T. E. & Goldsmith, G. R. The value of wet leaves. N. Phytol. 219, 1156–1169 (2018).
Feng, T. et al. Dew formation reduction in global warming experiments and the potential consequences. J. Hydrol. 593, 125819 (2021).
Gerlein-Safdi, C. et al. Dew deposition suppresses transpiration and carbon uptake in leaves. Agric. For. Meteorol. 259, 305–316 (2018).
Tomaszkiewicz, M., Abou Najm, M., Beysens, D., Alameddine, I. & El-Fadel, M. Dew as a sustainable non-conventional water resource: a critical review. Environ. Rev. 23, 425–442 (2015).
Fessehaye, M. et al. Fog-water collection for community use. Renew. Sustain. Energy Rev. 29, 52–62 (2014).
Kidron, G. J. Angle and aspect dependent dew and fog precipitation in the Negev desert. J. Hydrol. 301, 66–74 (2005).
Chiodi, A. M., Potter, B. E. & Larkin, N. K. Multi-decadal change in western US nighttime vapor pressure deficit. Geophys. Res. Lett. 48, e2021GL092830 (2021).
Tomaszkiewicz, M. et al. Projected climate change impacts upon dew yield in the Mediterranean basin. Sci. Total Environ. 566, 1339–1348 (2016).
Walker, B. H., Ludwig, D., Holling, C. S. & Peterman, R. N. Stability of semi-arid savanna grazing systems. J. Ecol. 69, 473–498 (1981).
Schlesinger, W. H. et al. Biological feedbacks in global desertification. Science 247, 1043–1048 (1990).
D’Odorico, P., Bhattachan, A., Davis, K., Ravi, S. & Runyan, C. Global desertification: drivers and feedbacks. Adv. Water Res. 51, 326–344 (2013).
Reynolds, J. F. et al. Global desertification: building a science for dryland development. Science 316, 847–851 (2007). Highlights the loss of ecosystem services as a result of dryland desertification.
Eldridge, D. J. et al. Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722 (2011). Provides a compehenseive analysis of the shrub enrochment effects on dryland functions.
IPCC Special Report on Climate Change and Land (eds Shukla, P. R. et al.) (IPCC, 2019).
Yang, H. et al. Tropical expansion driven by poleward advancing midlatitude meridional temperature gradients. J. Geophys. Res. Atmos. 125, e2020JD033158 (2020).
Berghuijs, W. R., Woods, R. A. & Hrachowitz, M. A precipitation shift from snow towards rain leads to a decrease in streamflow. Nat. Clim. Change 4, 583–586 (2014).
Ayyad, M. A., Fakhry, A. M. & Moustafa, A.-R. A. Plant biodiversity in the Saint Catherine area of the Sinai peninsula. Egypt. Biodivers. Conserv. 9, 265–281 (2000).
Global Land Outlook 2017 (UNCCD, 2017).
Van Ittersum, M. K. et al. Can sub-Saharan Africa feed itself? Proc. Natl Acad. Sci. USA 113, 14964–14969 (2016).
Redo, D., Aide, T. M. & Clark, M. L. Vegetation change in Brazil’s dryland ecoregions and the relationship to crop production and environmental factors: Cerrado, Caatinga, and Mato Grosso, 2001–2009. J. Land Use Sci. 8, 123–153 (2013).
Meyfroidt, P., Lambin, E. F., Erb, K.-H. & Hertel, T. W. Globalization of land use: distant drivers of land change and geographic displacement of land use. Curr. Opin. Environ. Sustain. 5, 438–444 (2013).
Rulli, M. C., Saviori, A. & D’Odorico, P. Global land and water grabbing. Proc. Natl Acad. Sci. USA 110, 892–897 (2013).
Müller, M. F. et al. Impact of transnational land acquisitions on local food security and dietary diversity. Proc. Natl Acad. Sci. USA 118, e2020535118 (2021).
Chiarelli, D. D. et al. Competition for water induced by transnational land acquisitions for agriculture. Nat. Commun. 13, 505 (2022).
Dell’Angelo, J., D’Odorico, P., Rulli, M. C. & Marchand, P. The tragedy of the grabbed commons: coercion and dispossession in the global land rush. World Dev. 92, 1–12 (2017).
Rosa, L. et al. Potential for sustainable irrigation expansion in a 3 °C warmer climate. Proc. Natl Acad. Sci. USA 117, 29526–29534 (2020).
Wang, L. & D’Odorico, P. The limits of water pumps. Science 321, 36–37 (2008).
OECD-FAO Agricultural Outlook 2021–2030 (OECD and FAO, 2021).
Qi, J., Xin, X., John, R., Groisman, P. & Chen, J. Understanding livestock production and sustainability of grassland ecosystems in the Asian Dryland Belt. Ecol. Process. 6, 22 (2017).
Godde, C. M. et al. Global rangeland production systems and livelihoods at threat under climate change and variability. Environ. Res. Lett. 15, 044021 (2020).
Herrero, M. et al. Exploring future changes in smallholder farming systems by linking socio-economic scenarios with regional and household models. Glob. Environ. Change 24, 165–182 (2014).
Bannari, A., Morin, D., Bonn, F. & Huete, A. A review of vegetation indices. Remote Sens. Rev. 13, 95–120 (1995).
Qiu, B. et al. Dense canopies browning overshadowed by global greening dominant in sparse canopies. Sci. Total Environ. 826, 154222 (2022).
Burrell, A. L., Evans, J. P. & Liu, Y. Detecting dryland degradation using time series segmentation and residual trend analysis (TSS-RESTREND). Remote Sens. Environ. 197, 43–57 (2017).
Bastin, J.-F. et al. The extent of forest in dryland biomes. Science 356, 635–638 (2017).
Griffith, D. M. et al. Comment on ‘The extent of forest in dryland biomes’. Science 358, eaao1309 (2017).
Teckentrup, L. et al. Assessing the representation of the Australian carbon cycle in global vegetation models. Biogeosciences 18, 5639–5668 (2021).
MacBean, N. et al. Dynamic global vegetation models underestimate net CO2 flux mean and inter-annual variability in dryland ecosystems. Environ. Res. Lett. 16, 094023 (2021). Highlights the often-neglected uncertainties in the prediction of dryland productivity.
Paschalis, A. et al. Rainfall manipulation experiments as simulated by terrestrial biosphere models: where do we stand? Glob. Change Biol. 26, 3336–3355 (2020).
Whitley, R. et al. A model inter-comparison study to examine limiting factors in modelling Australian tropical savannas. Biogeosciences 13, 3245–3265 (2016).
Hartley, A. J., MacBean, N., Georgievski, G. & Bontemps, S. Uncertainty in plant functional type distributions and its impact on land surface models. Remote Sens. Environ. 203, 71–89 (2017).
MacBean, N. et al. Testing water fluxes and storage from two hydrology configurations within the ORCHIDEE land surface model across US semi-arid sites. Hydrol. Earth Syst. Sci. 24, 5203–5230 (2020).
Burrell, A., Evans, J., De & Kauwe, M. Anthropogenic climate change has driven over 5 million km2 of drylands towards desertification. Nat. Commun. 11, 3853 (2020).
De Kauwe, M. G., Medlyn, B. E. & Tissue, D. T. To what extent can rising [CO2] ameliorate plant drought stress? N. Phytol. 231, 2118–2124 (2021).
Zhu, Z. et al. Greening of the Earth and its drivers. Nat. Clim. Change 6, 791–795 (2016).
Bernacchi, C. J. & VanLoocke, A. Terrestrial ecosystems in a changing environment: a dominant role for water. Annu. Rev. Plant Biol. 66, 599–622 (2015).
Roderick, M. L., Greve, P. & Farquhar, G. D. On the assessment of aridity with changes in atmospheric CO2. Water Resour. Res. 51, 5450–5463 (2015).
Anderegg, W. R., Trugman, A. T., Bowling, D. R., Salvucci, G. & Tuttle, S. E. Plant functional traits and climate influence drought intensification and land–atmosphere feedbacks. Proc. Natl Acad. Sci. USA 116, 14071–14076 (2019).
Zhou, S. et al. Land–atmosphere feedbacks exacerbate concurrent soil drought and atmospheric aridity. Proc. Natl Acad. Sci. USA 116, 18848–18853 (2019).
Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Change 3, 52–58 (2013).
Abdelmoaty, H. M., Papalexiou, S. M., Rajulapati, C. R. & AghaKouchak, A. Biases beyond the mean in CMIP6 extreme precipitation: a global investigation. Earth’s Future 9, e2021EF002196 (2021).
Dunkerley, D. L. Light and low-intensity rainfalls: a review of their classification, occurrence, and importance in landsurface, ecological and environmental processes. Earth Sci. Rev. 214, 103529 (2021).
Zhu, Y. & Yang, S. Interdecadal and interannual evolution characteristics of the global surface precipitation anomaly shown by CMIP5 and CMIP6 models. Int. J. Climatol. 41, E1100–E1118 (2021).
Cuthbert, M. O. et al. Observed controls on resilience of groundwater to climate variability in sub-Saharan Africa. Nature 572, 230–234 (2019).
Miguez-Macho, G. & Fan, Y. Spatiotemporal origin of soil water taken up by vegetation. Nature 598, 624–628 (2021).
Potapov, P. et al. Global maps of cropland extent and change show accelerated cropland expansion in the twenty-first century. Nat. Food 3, 19–28 (2022).
Trabucco, A. & Zomer, R. Global aridity index and potential evapotranspiration (ET0) climate database v.2. Figshare https://doi.org/10.6084/m9.figshare.7504448.v4 (2019).
Paschalis, A., Fatichi, S., Katul, G. G. & Ivanov, V. Y. Cross-scale impact of climate temporal variability on ecosystem water and carbon fluxes. J. Geophys. Res. Biogeosci. 120, 1716–1740 (2015).
Acknowledgements
We acknowledge support from Division of Earth Sciences of National Science Foundation (EAR‐1554894). N.M. acknowledges funding from NASA Carbon Cycle Program grant no. 80NSSC21K1709, S.M. from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant no. 101001608), G.V. from European Commission and Swedish Research Council for Sustainable Development FORMAS (grant 2018-02787) for funding in the frame of the collaborative international consortium iAqueduct, financed under the 2018 Joint call, and M.C.R. from the EU PRIMA Programme under Horizon 2020 European Union’s Framework Programme for Research and Innovation (NEXUS-NESS an Art.185 initiative grant no, 2042). We thank the TRENDY v.7 modellers for providing simulations and the AmeriFlux, OzFlux and FLUXNET site principal investigators for providing the in situ eddy covariance flux tower CO2 and ET fluxes used to produce Fig. 6.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Climate Change thanks Arden Burrell, Akash Koppa and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Notes 1–3, Fig. 1, Table 1 and references.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wang, L., Jiao, W., MacBean, N. et al. Dryland productivity under a changing climate. Nat. Clim. Chang. 12, 981–994 (2022). https://doi.org/10.1038/s41558-022-01499-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41558-022-01499-y
This article is cited by
-
Dryland hydroclimatic response to large tropical volcanic eruptions during the last millennium
npj Climate and Atmospheric Science (2024)
-
Survival and rapid resuscitation permit limited productivity in desert microbial communities
Nature Communications (2024)
-
Plant responses to changing rainfall frequency and intensity
Nature Reviews Earth & Environment (2024)
-
Streamflow decreases in response to acid deposition in a subtropical forest watershed in China
Communications Earth & Environment (2023)
-
Response of water-use efficiency to phenology in the natural forest and grassland of the Loess Plateau in China
Science China Earth Sciences (2023)
