
Abstract
Uncoupling protein-3 (UCP-3) is a recently identified member of the mitochondrial transporter superfamily1,2 that is expressed predominantly in skeletal muscle1,2. However, its close relative UCP-1 is expressed exclusively in brown adipose tissue, a tissue whose main function is fat combustion and thermogenesis. Studies on the expression of UCP-3 in animals and humans in different physiological situations support a role for UCP-3 in energy balance and lipid metabolism3,4. However, direct evidence for these roles is lacking. Here we describe the creation of transgenic mice that overexpress human UCP-3 in skeletal muscle. These mice are hyperphagic but weigh less than their wild-type littermates. Magnetic resonance imaging shows a striking reduction in adipose tissue mass. The mice also exhibit lower fasting plasma glucose and insulin levels and an increased glucose clearance rate. This provides evidence that skeletal muscle UCP-3 has the potential to influence metabolic rate and glucose homeostasis in the whole animal.
Similar content being viewed by others

Main
The human α-skeletal actin promoter was used to drive tissue-directed expression of a human UCP-3 transgene in C57BL/6 × CBA mice ( Fig. 1a). This promoter is well characterized, and the 2.2-kb fragment used (a gift from R. Prinjha and F. S. Walsh) contains all the necessary elements for selective expression in skeletal muscle5. During development, α-cardiac actin is the predominant isoform of sarcomeric α-actin in mice6, and it is only post-partum that there is a switch to α-skeletal actin5. Thus, by using the α-skeletal actin promoter, any possibility that embryonic expression of UCP-3 might interfere with development was minimized. The UCP-3 overexpressing animals had normal gestation, birth and litter sizes, and they were viable and outwardly healthy. The mice were developmentally normal in that femur length was similar between groups ( 14.1 ± 0.2 mm in wild-type and 14.2 ± 0.1 mm in UCP-3 transgenic (tg) mice at 17 weeks of age; mean ± s.e.m. for 10 animals per group). A number of founders were generated and three independent lines were bred to homozygosity. Of these, two independent lines showed a significant reduction in body weight. The third expressed low levels of human UCP-3. The line expressing the highest levels of human UCP-3 was used to examine the phenotype further. Quantitative polymerase chain reaction with reverse transcription (RT–PCR) of messenger RNA for the human transgene in a single UCP-3tg mouse confirmed that expression was largely confined to skeletal muscle with little or no ectopic expression in other tissues (for example, stomach smooth muscle <1% of skeletal muscle) except brown adipose tissue (Fig. 1b). However, measurement of transgenic UCP-3 expression in brown adipose tissue in a larger sample (n = 10) showed that the expressed transgene was only 1% of that in skeletal muscle (Table 1). Total UCP-3 expression was increased 66-fold in skeletal muscle but by only 50% in brown adipose tissue (representing 3% of the levels of endogenous UCP-1). The phenotype described here is therefore due primarily to expression of the transgene in skeletal muscle. As expected, protein expression was confined to the mitochondrial fraction of transgenic muscle (Fig. 1c).

a, Map of the 3.9-kilobase (kb) construct used for microinjection of [C57BL/6 × CBA] F2 fertilized eggs. Hatched and stippled blocks: artificial intron and SV40 polyadenylation signal from pIRES plasmid, respectively. b, Quantitative RT–PCR (TaqMan, PE Biosystems) for human UCP-3. BAT and WAT, brown and white adipose tissue, respectively. c, Immunoblot of human UCP-3 expression at relative molecular mass 34,000 (M r 34K) in the mitochondrial fraction (m) of UCP-3tg mice, but not wild type, using a rabbit anti-human UCP-3 antibody (Alpha Diagnostic Inc). s, supernatant.
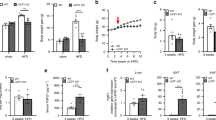
Although data on physiological measurements and blood analytes in this report pertain to males, the phenotype was also observed in female UCP-3tg mice. As UCP-3 overexpression in skeletal muscle was proposed to result in an increase in energy expenditure, it was not surprising that the predominant phenotypic characteristic was a reduction in body weight ( Fig. 2a). Placing the animals on an energetically equivalent, but palatable diet7 encouraged consumption (for example, 96.5 kJ per 24 h compared with 78.6 kJ per 24 h for UCP-3tg mice). It also accelerated weight gain in wild-type animals, but not in UCP-3tg animals: weight gain in UCP-3tg mice tended to plateau at the point of diet change. A surprising finding was that, in spite of their markedly lower body weight, UCP-3tg mice were hyperphagic. They consumed between 15% and 28% more food energy than wild-type mice on the normal diet from 4 to 8 weeks of age, and 33–54% more on the palatable diet between 8 and 12 weeks of age (Fig. 2b). Palatable diets evoke increased sympathetic nerve activity8,9,10, which stimulates thermogenesis in brown adipose tissue. This, together with the effects of UCP-3 overexpression, may account for the plateau of body weight gain despite increased energy intake. Moreover, further activation of transgenic UCP-3 by products of sympathetically mediated lipolysis (for example, fatty acids) cannot be ruled out. The difference in energy intake between transgenic mice and their wild-type littermates was maintained when the animals were returned to a normal diet. Despite a 50% increase in food consumption, plasma triglycerides and non-esterified fatty acids were similar between the UCP-3tg mice and wild-type controls, suggesting that fat combustion was higher in UCP-3tg mice (Table 1).

a, UCP-3tg mice (filled squares) weigh less than their wild-type littermates (open squares) when measured between 4 and 14 weeks of age. Shaded portion of the graph indicates a switch to palatable diet and coincides with a plateau in weight gain. b, Twenty-four-hour food intake in grams per mouse, other indices as for a. c, Oral glucose tolerance curve at the end of the initial period on normal diet (eight weeks). Data are mean ± s.e.m. (on some data points error bars are within symbols; asterisk, P < 0.05) for 12 animals per group.
Magnetic resonance imaging (MRI) analysis following four weeks on the palatable diet revealed a striking reduction in adipose tissue mass in UCP-3tg mice (Fig. 3a). A 44% and 57% decrease in the ratio of adipose tissue volume to total animal volume was seen in males and females, respectively (Fig. 3b). These alterations may be the predominant factor contributing to reduced weight in these animals. Despite the reduction in adipose tissue content of the UCP-3tg mice, plasma leptin levels were not significantly reduced (Table 1).

a, Abdominal images of wild-type and UCP-3tg mice following four weeks on palatable diet. b, Comparison between fat as a percentage of body volume in wild-type and UCP-3tg mice. Seven two-dimensional transverse sections for the anterior end of the left kidney through to the posterior end of the right kidney were used for the calculation. Graphs represent mean values of % fat to body volume ± s.e.m. for six animals per group. Statistical significance was calculated using Student's t-test.
The phenotype of the UCP-3tg mouse is consistent with an increase in energy expenditure, which was confirmed directly. Resting oxygen consumption was 25% higher with normal diet at 8 weeks of age (30.5 ± 2.2 ml per animal per hour versus 38.3 ± 1.6 ml per animal per hour in UCP-3tg mice; P < 0.05) and 40% with palatable diet at 12 weeks of age (39.8 ± 3.0 ml per animal per hour in wild-type versus 55.9 ± 4.0 ml per animal per hour in UCP3tg mice; P = 0.03). However, locomotor activity was not significantly increased (Table 1). If oxygen consumption is corrected by body weight, the increased consumption of the transgenic mice was 77% (P < 0.02) on normal diet and 91% ( P < 0.005) on palatable diet. Time spent on an accelerating Rotorod, as an index of muscle motor coordination, was also unaffected by UCP-3 overexpression (Table 1). Core temperature was not affected by the presence of the transgene (38.32 ± 0.07 °C in wild-type and 38.33 ± 0.13 °C in UCP-3tg mice), although muscle temperature was increased (37.52 ± 0.32 °C in wild-type mice versus 38.72 ± 0.38 °C in UCP-3tg; P < 0.05) at 14 weeks of age. The increased muscle temperature was similar to the increase in brown adipose tissue temperature elicited by β 3-adrenoceptor agonists in rats11. Rodents can sustain marked increases in metabolic rate, with little increase in the temperature of the heat-generating tissue, by rapidly dissipating heat to their environment by radiation and convection12.
An emerging hypothesis on the role of UCP-3 favours a primary role in lipid substrate utilization3,4. This model was proposed because UCP-3 mRNA levels change markedly, in parallel with altered fatty acid oxidation. For example, 24-hour fasting increases UCP-3 mRNA 6–12-fold, depending on muscle type, without overt changes in whole-body metabolic rate3,4. Thermogenesis is clearly occurring in UCP-3tg mice, but in this context it may be a by-product of greatly increased fatty acid oxidation. Similarly, it has been argued that UCP-1-mediated thermogenesis may be a by-product of fatty acid oxidation, so that animals can survive on a low protein diet without becoming obese13.
UCP-3tg mice also displayed reduced fasting plasma glucose levels, increased glucose clearance following an oral glucose load and reduced plasma insulin levels (Fig. 2c and Table 1), indicating that they may be more insulin-sensitive than their wild-type littermates. β 3-adrenoceptor agonists, which increase metabolic rate and thermogenesis, also elicit weight loss in rats by reducing white adipose tissue mass, and increase insulin sensitivity14,15,16.
We measured the respiration rate and membrane potential of isolated skeletal muscle mitochondria17 in six independent paired experiments. UCP-3 overexpression was associated with mitochondrial uncoupling, measured as a decrease in respiratory control ratio from 3.4 ± 0.1 to 2.4 ± 0.1 (P < 0.001). This was caused by a 26 ± 9% increase in state four respiration (using oligomycin to inhibit ATP synthesis, P < 0.003) from a control value of 109 ± 6 nmol O2 per minute per milligram protein, and by a 12 ± 6% decrease in state three respiration (P = 0.04). In parallel experiments (using nigericin to allow exchange of protons with potassium ions), the mitochondrial membrane potential of 168 ± 5 mV decreased by 12 ± 3% (P = 0.001). Increased state four respiration accompanied by a decreased membrane potential is diagnostic of mitochondrial uncoupling. However, changes in mitochondrial proton conductance following physiologically mediated changes in UCP-3 levels have not been described, and a mitochondrial transport function of UCP-3 cannot be ruled out.
Some studies indicate a role for UCP-3 in the regulation of energy expenditure in humans18,19, but there are also studies that fail to make an association20. The data here are clear, however: despite increased energy intake, overexpression of UCP-3 elicits a marked reduction in body weight, a marked reduction in adiposity and improved insulin sensitivity. Our data are not evidence that UCP-3 is involved in the aetiology of obesity, but they do suggest that enhancement of UCP-3 expression, or stimulation of its activity, is a promising approach for the treatment of this disease.
Methods
Animals
Mice were maintained in accordance with procedures outlined in the Home Office Animals (Scientific procedures) Act 1986. They were housed in groups of 12 (in cages of 3) and maintained on a 12-h light/12 hour dark cycle (lights on at 06:00 GMT). Normal diet was Teklad 2018 (13.7 kJ g-1) and the palatable diet (12.6 kJ g-1) was prepared as described7. The animals were implanted with Implantable Programmable Temperature Transponder chips (IPTT-100, Biomedic Data Systems Inc.) over the rear thigh muscles for identification and muscle temperature estimation.
Generating the transgenic mouse
The transgene fragment (Fig. 1a) was excised from its vector by restriction digestion and purified for injection (2 ng µl-1). Donor mice were prepared using standard procedures21. A Zeiss Axiovert 135M inverted microscope and Leica micromanipulators were used for the microinjection of fertilized eggs. The animals were maintained in a VAF+ animal house. The recipient eggs were [C57BL/6 × CBA] F2 hybrids.
RNA analysis and immunoblotting
Quantification of mRNA transcripts was performed using the PCR-based 5′ nuclease assay22 with gene-specific fluorogenic TaqMan hydrolysis probes23. Total RNA was treated with deoxyribonuclease I (Promega) and reverse transcribed using random nonamers (Stratagene) and MMLV reverse transcriptase (Life Technologies). After first-strand cDNA synthesis, transcript cDNAs were measured using TaqMan assay oligonucleotide primers and fluorogenic probes designed for human and mouse UCP-3 (amplicon 915–1011 of accession U84763 and amplicon 51–128 of accession AF032902, respectively) and mouse UCP-1 (amplicon 11–109 of accession U63419).
Immunoblots of human UCP-3 were carried out on mouse mitochondrial fractions24 and separated on SDS–PAGE using rabbit anti-human UCP-3 antibodies (Alpha Diagnostic Inc.), anti-rabbit IgG horse radish peroxidase secondary antibody (Amersham) and ECL detection kit (Pierce-Warriner).
Blood analytes
We measured leptin and insulin in plasma taken from 5-h fasted mice using commercially available ELISA (Crystal Chem Inc.). Plasma glucose, triglycerides, non-esterified fatty acids and total cholesterol were measured in the same samples spectrophotometrically using a Cobas Mira plus Clinical Chemistry Analyser (Roche Diagnostics).
Physiological measurements
For food intake measurements, mice were transferred to cages with grid bottoms for 24 h once a week. Body weight was also determined weekly. Oral glucose tolerance tests were performed at 8 weeks of age according to ref. 25. Oxygen consumption was measured using a Servomax oxygen analyser model 580A. Samples were monitored continuously in 20-min blocks with room air as reference. Mice were placed in the chamber during the light phase and the data collected for a 5-h period after the animals had settled into the new environment, but before activity increased owing to anticipation of the dark phase.
Magnetic resonance imaging analysis
Images were obtained on a Bruker AMX300 interfaced to a 18.3-cm 7-T magnet. Animals were anaesthetized using 5% isoflurane and maintained using 1.5% isoflurane and 100% O2. We monitored heart rate and respiration using an ECG and a fibre-optic mechanical probe, respectively. Data acquisitions were triggered to the flat part of the respiratory cycle. A three-dimensional multi-echo sequence was used, acquiring a field of view of 6 × 6 × 6 cm and a matrix size of 128 ± 96 ± 96. The experimental time was 38 min using 8 echoes with an echo time of 6.6 ms, a repetition time of ∼2 s and a spectral width of 100 kHz. The data set was zero filled to 256 × 128 × 128 for reconstruction.
All three-dimensional images were quantified using seven sequential transverse sections, 2 mm apart. The total area quantified and the anatomical boundaries were consistent between each animal and the fat depots within each mouse were quantified using the standard software tools available on ParaVision 2.0 (Bruker).
Activity measurements
Locomotor activity was tested in unhabituated mice as described26. Mice (one per cage) were placed in the activity monitor for 24 h, with food and water available ad lib. Locomotor activity was recorded automatically every 30 min. Rotorod experiments were conducted as described27.
References
Boss, O. et al. Uncoupling protein-3, a new member of the mitochondrial carrier family with tissue-specific expression. FEBS Lett. 408, 39–42 (1999).
Vidal-Puig, A., Solanes, G., Grujic, D., Flier, J. S. & Lowell, B. B. UCP3—an uncoupling protein homolog expressed preferentially and abundantly in skeletal-muscle and brown adipose-tissue. Biochem. Biophys. Res. Commun. 235, 79– 82 (1997).
Samec, S., Seydoux, J., Dulloo, A. G. Role of UCP homologs in skeletal-muscles and brown adipose-tissue—mediators of thermogenesis or regulators of lipids as fuel substrate. FASEB J. 12, 715–724 ( 1998).
Samec, S., Seydoux, J., Dulloo, A. G. Post-starvation gene expression of skeletal muscle uncoupling protein 2 and uncoupling protein 3 in response to dietary fat levels and fatty acid composition—A link with insulin resistance. Diabetes 48, 436–441 ( 1999).
Brennan, K. J. & Hardeman, E. C. Quantitative analysis of the human alpha-skeletal actin gene in transgenic mice. J. Biol. Chem. 268, 719–725 ( 1993).
Grunning, P. et al. Differential patterns of transcript accumulation during human myogenesis. Mol. Cell. Biol. 7, 4100– 4114 (1987).
Widdowson, P. S., Upton, R., Buckingham, R., Arch, J. & Williams, G. Inhibition of food response to intracerebroventricular injection of leptin is attenuated in rats with diet-induced obesity. Diabetes 46, 1782– 1785 (1997).
Bellisle, F., Louis-Sylvestre, J., Demozay, F., Blazy, D. & Le Magnen, J. Cephalic phase of insulin secretion and food stimulation in humans, a new perspective. Am. J. Physiol. 249, E639–E645 ( 1985).
Leblanc, J. & Brondel, L. Role of palatability on meal-induced thermogenesis in human subjects. Am. J. Physiol. 248 , E333–E336 (1985).
Diamond, P., Brodel, L. & LeBlanc, J. Palatability and postprandial thermogenesis in dogs. Am. J. Physiol. 248, E75– E79 (1985).
Arch, J. R. et al. Treatment of obesity with thermogenic beta-adrenoceptor agonists, studies on BRL 26830A in rodents. Int. J. Obes. 8 (suppl. 1), 1–11 (1984).
Gordon, C. J. Temperature Regulation in Laboratory Rodents (Cambridge Univ. Press, 1993).
Stock, M. J. Gluttony and thermogenesis revisited. Int. J. Obes. 23, 1105–1117 (1999).
Largis, E. E., Burns, M. G., Muenkel, H. A., Dolan, J. A. & Claus, T. H. Antidiabetic and antiobesity effects of a highly selective β3-adrenoceptor agonist (CL 316,243). Drug Dev. Res. 32, 69–76 (1994).
Weyer, C., Tataranni, A., Snitker, S., Danforth, E. & Ravussin, E. Increase in insulin action and fat oxidation after treatment with CL 316,243, a highly selective beta3-adrenoceptor agonist in humans. Diabetes 47, 1555– 1561 (1998).
Liu, X., Perusse, F. & Bukoweicki, L. J. Mechanisms of the antidiabetic effects of the beta 3-adrenergic agonist CL-316243 in obese Zucker-ZDF rats. Am. J. Physiol. 274, R1212–R1219 (1998).
Cadenas, S. et al. UCP2 and UCP3 rise in starved rat skeletal muscle but mitochondrial proton conductance is unchanged. FEBS Lett. 462, 257–260 (1999).
Argyropoulos, G. et al. Effects of mutations in the human uncoupling protein 3 gene on the respiratory quotient and fat oxidation in severe obesity and type 2 diabetes. J. Clin. Invest. 102, 1345– 1351 (1998).
Otabe, S. et al. A genetic variation in the 5′ flanking region of the UCP3 gene is associated with body mass index in humans in interaction with physical activity. Diabetologia 43, 245– 249 (2000).
Chung, W. K. et al. the long isoform uncoupling protein-3 (UCP3L) in human energy homeostasis. Int. J. Obes. 23, S49– S50 (1999).
Hogan, B., Lacy, E., Beddington, R. & Constantini, F. Manipulating the Mouse Embryo 2nd edn (Cold Spring Harbor Press, 1994).
Holland, P. M., Abramson, R. D., Watson, R. & Gelfand, D. H. Detection of specific polymerase chain reaction product by utilizing the 5′-3′exonuclease activity of Thermus aquaticus DNA polymerase. Proc. Natl Acad. Sci. USA 88, 7276–7280 (1991).
Livak, K. J. Flood, S. J., Marmaro, J., Giusti, W. & Deetz, K. Oligonucleotides with fluorescent dyes at opposite ends provide a quenched probe system useful for detecting PCR product and nucleic acid hybridization. PCR Methods Appl. 4, 357–362 (1995).
Ernster, L. & Nordenbrand, K. Skeletal muscle mitochondria. Methods Enzymol. 10, 86– 94 (1967).
Young, P. W. et al. Repeat treatment of obese mice with BRL 49653, a new potent insulin sensitizer, enhances insulin action in white adipocytes. Association with increased insulin binding and cell-surface GLUT4 as measured by photoaffinity labeling. Diabetes 44, 1087– 1092 (1995).
Rogers, D. C. et al. Use of SHIRPA and discrimainant analysis to characterise marked differences in the behavioural phenotype of six inbred mouse strains. Behav. Brain. Res. 105, 207–217 (1999).
Jones, B. J. & Roberts, D. J. The quantitative measurement of motor inco-ordination in naive mice using an accelerating rotarod. J. Pharm. Pharmacol. 20, 302–304 (1968).
Acknowledgements
S.C. was supported by a Marie Curie (EC) fellowship.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Clapham, J., Arch, J., Chapman, H. et al. Mice overexpressing human uncoupling protein-3 in skeletal muscle are hyperphagic and lean. Nature 406, 415–418 (2000). https://doi.org/10.1038/35019082
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1038/35019082
This article is cited by
-
A transdermal treatment with MC903 ameliorates diet-induced obesity by reducing visceral fat and increasing myofiber thickness and energy consumption in mice
Nutrition & Metabolism (2023)
-
CD47 antisense oligonucleotide treatment attenuates obesity and its-associated metabolic dysfunction
Scientific Reports (2023)
-
Early high-fat feeding improves histone modifications of skeletal muscle at middle-age in mice
Laboratory Animal Research (2020)
-
Ccr7 null mice are protected against diet-induced obesity via Ucp1 upregulation and enhanced energy expenditure
Nutrition & Metabolism (2019)
-
Cytosolic lipid excess-induced mitochondrial dysfunction is the cause or effect of high fat diet-induced skeletal muscle insulin resistance: a molecular insight
Molecular Biology Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
