
Abstract
Interactions between microtubules and actin are a basic phenomenon that underlies many fundamental processes in which dynamic cellular asymmetries need to be established and maintained. These are processes as diverse as cell motility, neuronal pathfinding, cellular wound healing, cell division and cortical flow. Microtubules and actin exhibit two mechanistic classes of interactions — regulatory and structural. These interactions comprise at least three conserved 'mechanochemical activity modules' that perform similar roles in these diverse cell functions.
Similar content being viewed by others

Main
Over the past 35 years, great progress has been made towards understanding the roles of the microtubule and actin cytoskeletal filament systems in mechanical cellular processes such as dynamic shape change, shape maintenance and intracellular organelle movement. These functions are attributed to the ability of polarized cytoskeletal polymers to assemble and disassemble rapidly, and to interact with binding proteins and molecular motors that mediate their regulated movement and/or assembly into higher order structures, such as radial arrays or bundles. This allows, for example, microtubules to form a bipolar spindle that can move chromosomes into two daughter cells with high fidelity, and actin to mediate muscle contraction or promote protrusion at the leading edge of a migrating cell.
Although it is certainly true that microtubules and actin have such distinct roles, it has been evident for some time that interactions between these seemingly distinct filament systems exist. Vasiliev1 hinted at this years ago when he showed that an intact microtubule cytoskeleton was required to maintain the polarized distribution of actin-dependent protrusions at the leading edge of a migrating fibroblast. This suggested that the microtubule cytoskeleton somehow directs proper placement of actin polymerization- and contraction-based activities.
Since then, it has become clear that similar microtubule/actin interactions are a basic phenomenon that underlie many fundamental cellular processes, including cell motility, growth cone guidance, cell division, wound healing and cortical flow. In general, such cytoskeletal crosstalk occurs in processes that require dynamic cellular asymmetries to be established or maintained to allow rapid intracellular reorganization or changes in shape or direction in response to stimuli. Furthermore, the widespread occurrence of these interactions underscores their importance for life, as they occur in diverse cell types including epithelia, neurons, fibroblasts, oocytes and early embryos, and across species from yeast to humans. Thus, defining the mechanisms by which actin and microtubules interact is key to understanding a basic organizing principle for dynamic morphogenesis, which, in turn, is a step towards understanding health-related processes such as cancer, wound healing and neuronal regeneration. Recent investigations that shed light on these elusive interactions shall be the focus of our review.
'Structural' versus 'regulatory' interactions
What are the cellular and molecular bases of microtubule–actin cooperation? One popular viewpoint is the 'tensegrity model2,3, in which actomyosin generates tension against stiff microtubule 'struts' and adhesions to the substrate to stabilize or change cell shape. Although these principles may be applicable, we propose an alternative, not necessarily exclusive, hypothesis, in which the interactions between actin and microtubules may be classified as either 'regulatory' or 'structural'.
Regulatory interactions are those in which the two systems indirectly control each other through their effects on signalling cascades (Fig. 1a). The best understood example of regulatory interactions is provided by the Rho family of small GTPases, which regulate both microtubules and actin4. For example, RhoA mediates formation of contractile actin structures, such as stress fibres5, and at the same time promotes stabilization of a sub-population of microtubules6. Two key factors are known to function downstream of RhoA: Rho kinase, which promotes contractility by increasing phosphorylation of the regulatory light chain of myosin-2 (ref. 7), and the formin, mDia, which regulates actin polymerization into bundles8,9 and also mediates microtubule stabilization10. Similarly, Rac1 activity regulates the polymerization of both actin and microtubules to promote lamellipodial protrusion11,12. Rac1 controls actin through both Scar/WASP proteins that activate Arp2/3-dependent assembly of dendritic meshworks and Pak kinases that may regulate leading-edge actin treadmilling through ADF/cofilin proteins7,11,12. Pak kinases downstream of Rac1 also promote microtubule growth, probably by regulating the microtubule destabilizing protein, Op18/stathmin12,13.

(a) Rho GTPases and cytoskeletal filaments can regulate each other. Microtubule disassembly stimulates RhoA activity (left). Rho stabilizes microtubules through the formin mDia and also results in actomyosin contraction through stimulation of Rho kinase, resulting in myosin light chain phosphorylation. Microtubule growth stimulates Rac1 activity (right), which mediates actin polymerization and lamellipodial protrusion, and promotes further microtubule growth through activation of the Pak1–Op18 signalling pathway to generate a positive feedback loop. (b) Actin (red) and microtubules (green) can exhibit static or dynamic interactions. Interaction 1 shows a protein that possesses both actin- and microtubule-binding sites and could provide a static crosslink between the two polymers, as hypothesized for MAP2c. Interaction 2 shows a complex between an actin-based motor (blue) and a microtubule-based motor (orange), whereas interaction 3 shows a complex between a motor (yellow) and a binding protein (pink). Both types of interaction could move actin and microtubules relative to one another, as hypothesized for myosin V and kinesin, or myosin VI and Drosophila CLIP-190, respectively (see Table 1).
In turn, the activity of Rho proteins is regulated by microtubules and actin. Microtubule or actin disassembly activates RhoA14, whereas microtubule assembly promotes Rac1 activation15. Depolymerizing microtubules are thought to release the microtubule-bound Rho guanine nucleotide-exchange factor (GEF) GEF-H1 to activate RhoA16, whereas the mechanism through which microtubule growth activates Rac1 is completely unknown. Whether the local microtubule assembly state affects the local activation state of Rho GTPases within living cells awaits a direct demonstration.
Structural interactions are those in which actin and microtubules are physically linked (Fig. 1b). Experiments in which microtubules and actin filaments were imaged simultaneously in Xenopus laevis egg extracts indicate that there are both static and dynamic structural interactions17. Static interactions could be mediated by complexes between microtubule- and actin-binding proteins, or by individual proteins that can bind both filaments simultaneously. In contrast, dynamic interactions, in which there is relative movement between the two polymers, could involve a microtubule- or actin-based motor and an actin- or microtubule-binding protein, or combinations thereof. Although interesting, we will not discuss the switching of cellular cargo between microtubule- and actin-based motors in this review.
Identifying the molecules that mediate structural interactions between microtubules and actin is an important challenge for the future. There are several candidate proteins that contain homology to known actin- and microtubule-binding motifs or which localize to either filament networks under certain conditions (Table 1), but rigorous analysis of their microtubule/actin-crosslinking activity has not been performed. Although this review is not focused on intermediate filaments, they may be critical in mediating structural interactions between microtubules and actin, functioning through a family of cytoskeletal crosslinkers, the plakins. Plakins are large (relative molecular mass (Mr) 200–600K) multidomain proteins that bind intermediate filaments and are essential for maintaining tissue integrity18. Several plakins, including plectin, bullous pemphogoid antigen 1 (BPAG1) and microtubule-actin crosslinking factor (MACF), also contain actin- or microtubule-binding sites, and thus could potentially interlink all three filament systems18,19.
What evidence is there that 'structural interactions' between microtubules and actin actually occur in living cells? Such interactions have been difficult to detect, presumably because they are dynamic or transient. Furthermore, both polymers are so abundant that it is difficult in static images to differentiate filaments that just happen to cross over from those that are specifically bound. The strongest support for the existence of structural interactions in vivo comes from recent live-cell imaging studies, in which co-transport of both filaments at the same velocity and trajectory throughout the cell is taken as evidence for a binding interaction. This has been made possible by the use of multi-spectral time-lapse fluorescence microscopy and fluorescent speckle microscopy (FSM) of fiduciary-marked actin and microtubules labelled with spectrally distinct fluorophores20,21,22,23. The observation that microtubules and actin are efficiently co-transported (and so might structurally interact) was established in an in vitro assay in which microtubules moving on coverslip-bound motors pulled along actin bundles only in the presence of cytosolic factors17. Furthermore, in migrating cells, cotransport of microtubules and F-actin is regionally regulated and does not occur in the lamellipodium where the actin density is highest, arguing against the idea that microtubules might be non-specifically trapped by a dense moving actin meshwork20.
Microtubule–actin interactions in directed cell migration
The polarity of a migrating cell with a ruffling leading lamellipodium and contractile tail is reflected by the polarized organization of the underlying cytoskeleton (Fig. 2a). Microtubules in vertebrate tissue cells are arranged with their minus ends near the cell centre or anchored at the centrosome, which is often positioned between the nucleus and the leading edge. Microtubule plus ends radiate primarily towards the leading edge, where they display dynamic instability4. F-actin is arranged with the plus ('barbed') ends just subadjacent to the plasma membrane and the minus ('pointed') ends pointing towards the cell interior. At the leading edge are lamellipodia, in which actin forms a dendritic network, and filopodia, which are comprised of parallel actin bundles11. Actin polymerization from the plus ends at the leading edge of lamellipodia and filopodia is balanced by a myosin-powered, rearward movement of the lamellum actin meshwork known as retrograde flow24. Behind the lamellum, actin bundles and meshwork move towards the cell front to create a 'convergence zone' where retrograde and anterograde actin motions meet20,25. Here, myosin-2 is concentrated, suggesting high contractility26, and actin filament disassembly is prominent20. To move, the cell generates traction through contractile actin bundles pulling on focal contacts attached to the substrate, which must be assembled at the cell front and disassembled at the cell rear27.

(a) In migrating tissue cells, microtubule (green) minus ends are organized by the centrosome, which is positioned between the nucleus and the leading edge, and their plus ends are orientated towards the leading edge plasma membrane. In lamellipodia (LA), F-actin (red) is in a meshwork that undergoes retrograde flow towards the convergence zone (CZ), where myosin (blue dots) is concentrated and contractility is high. Stress fibres are contractile actomyosin bundles with their ends anchored in focal adhesions. Insets show putative microtubule–actin interactions in different regions of a migrating cell. In the lamellum (panel 1), microtubules are coupled to F-actin undergoing retrograde flow. This process compresses and breaks microtubules to promote regional microtubule turnover. Microtubules could target focal adhesions (panel 2) by crosslinking to and growing along focal-adhesion-associated actin bundles. Microtubule plus ends may be anchored at the cell cortex (panel 3) through interactions between plus-end-binding proteins and actin-binding proteins to orientate the MTOC towards the direction of migration. (b) Neuronal growth cone. The growth cone has two kinetically distinct zones of actin translocation associated with filopodial (red) structures and actin meshwork (black) in the periphery, and actin arcs (blue) in the transition zone. Both F-actin filopodia and actin arcs guide the assembly and transport of microtubules. Dynamic unbundled microtubules (green) polymerize into the periphery along filopodia and are simultaneously cleared from the periphery by depolymerization or coupling to retrograde actin flow. Microtubules caught in retrograde flow can also bend and break, exposing unstable minus ends and new plus ends. This allows microtubule turnover within growth cones. Less dynamic microtubules (brown) extending along the lateral sides of the growth cone are packed into the central domain (light brown microtubules) by coupling to transverse actin arc movements. P (light blue), T (dark blue), and C (brown) refer to the peripheral lamellar actin domain, transition zone and central domain, respectively. (c) A region at the edge of a wound in the plasma membrane of a Xenopus oocyte. F-actin (red lines) and myosin-2 (blue dots) concentrate at wound borders as a result of flow from the outlying cortex (arrows) and local assembly (short red lines and dots). Microtubules (green lines) are pulled towards the wound border by flowing F-actin and are also assembled near the wound border (short green lines and dots). As microtubules flow towards the contractile wound border, they are buckled and broken.
Recent in vivo and in vitro studies have shown that actin has a major influence on the organization of microtubules in migrating cells. This was first demonstrated by the observation that microtubules are transported rearward in the lamella of motile cells and that this depends on actomyosin28,29,30. Labelling of actin and microtubules with spectrally distinct fluorophores and analysis by dual-wavelength FSM showed that microtubules are coupled to actin retrograde flow in the lamella, and to anterograde motion of actin in the cell body (Fig. 2a, panel 1, and Fig. 3a)20,26. Microtubules also grow along actin bundles (Fig. 2a, panel 2) and microtubule ends are often dragged through the cytoplasm by their connection to moving actin bundles20.

Migrating newt lung epithelial cells (a), growth cones from Aplysia bag cell neurons (b) and a wound edge in a Xenopus oocyte (c) are shown for comparison. Cells were comicroinjected with green fluorescent tubulin and red fluorescent actin (a) or phalloidin (b, c) and imaged by dual-wavelength fluorescent speckle (FSM; a, b) or confocal microscopy (c). In time-lapse microscopy, the simultaneous movement of actin and microtubules at the same trajectory and velocity is taken as evidence for structural interactions between the two polymers. (a) In newt lung epithelial cells, the movements of microtubules and F-actin are coupled in the lamellum. The top panel shows a single FSM image, the leading edge is at the top. The bottom panel shows a time montage of the boxed region in the upper panel. This microtubule is transported rearward while simultaneously growing towards the leading edge. The white horizontal line tracks the retrograde movement of a speckle on the microtubule, which is moving at the same velocity as immediately adjacent speckles in the lamella actin meshwork. Frames are at 10-s intervals. Scale bar represents 10 μm in both panels. (b) A single FSM image (top), showing the co-alignment of filopodial F-actin bundles (red) and microtubules (green) in the periphery of an Aplysia bag cell growth cone. Same frame showing the overlay of microtubules with the DIC channel (middle). Dimensions for top and middle images are 24 × 38 μm. A timelapse montage (bottom), showing an example of microtubule alignment along a filopodial actin bundle. Initially, the microtubule is uncoupled from the flow and straightens out along the filopodium (blue circles). The microtubule then begins translocating at the same rate as adjacent actin speckles, where it buckles and forms a loop (blue triangle). Frames are at 12-s intervals. Dimensions for each frame are 19 × 5 μm. (c) Micrograph of wound edge in a Xenopus oocyte (top). Microtubules (green) flow towards the wound edge surrounded by a contractile actomyosin ring (actin, red). A timelapse montage (bottom), in which microtubules and actin motion is coupled. Colocalization is shown in yellow. At the same time that microtubules are transported towards the wound edge, they polymerize and grow away from the edge. Frames are at 15-s intervals.
Several non-exclusive hypotheses have been proposed to explain how cells use structural and regulatory actin–microtubule interactions to generate movement. One hypothesis is that cell motility depends on the structural linkage of microtubules to actin retrograde flow, which in turn establishes and maintains a regulatory Rho GTPase signalling gradient that perpetuates motility4. The linkage of microtubules to actin retrograde flow requires a compensatory net growth of microtubules towards the leading edge. Behind the lamellum in the convergence zone, microtubule breakage and depolymerization occurs as a result of the compressive forces of the converging actin to which they are bound20,26,28. Thus, microtubule linkage to regional actin movements results in a gradient of microtubule assembly states in the cell with plus-end growth at the leading edge and minus-end shortening predominating behind the lamellum28,31. This could, in turn, create regional regulatory interactions between microtubules and actin. Microtubule growth could promote local activity of Rac in the cell front to drive lamellipodial protrusion, focal complex formation and perpetuate further microtubule growth12,15. Microtubule shortening could activate RhoA behind the lamellum to drive actomyosin contraction and promote the stabilization of a sub-population of microtubules, possibly to protect them from breakage and thus maintain the overall polarization of the microtubule cytoskeleton6,14. In support of this hypothesis, FRET-based visualization of Rac1 activity has revealed a gradient from the leading edge32,33, although RhoA activity has not yet been visualized. The adenomatous polyposis coli (APC) protein is one candidate that might mediate the activation of Rac1 driven by microtubule growth in migrating cells. APC localizes to growing microtubule plus ends in ruffling cell protrusions34,35 and binds Asef, a Rac1-specific GEF that stimulates lamellipodia formation and cell migration36,37. An equally interesting candidate is IQGAP1, which binds Rac, Cdc42 and actin, and also associates with growing microtubule plus ends through CLIP-170 (ref. 38). Furthermore, disruption of the interaction between IQGAP1, Rac1 and Cdc42 delocalizes lamellipodial activity38.
A second hypothesis is that microtubule-actin interactions orientate towards the leading edge, which could then direct the delivery of signalling molecules or membrane components required for lamellipodial protrusion39. Recent studies have shown that microtubule organizing centre (MTOC) re-orientation during cell migration is mediated by the Rho GTPase Cdc42 and the microtubule motor, cytoplasmic dynein40,41. It has been suggested that microtubule ends are 'captured' by structural crosslinks to specific sites in the actin cortex whose location is defined by Cdc42, and that there, the motor activity of cytoplasmic dynein may pull the MTOC in front of the nucleus39 (Fig. 2a, panel 3). A model for this comes from cell division in Saccharomyces cerevisiae. Here, a spindle microtubule is guided to the bud site by a microtubule plus-end complex consisting of Kar9, the Class V myosin motor Myo2 and Bim1 (ref. 42). Subsequently, the microtubule becomes captured at a Cdc42-defined site in the actin-rich cortical bud tip of the daughter cell, where cytoplasmic dynein pulls the mitotic spindle into the daughter cell39,43. A similar mechanism may exist for anchoring microtubules in migrating cells, as EB1 and APC (putative homologues of Bim1 and Kar9, respectively) both localize to growing microtubule ends44,45. Dynein is also well positioned to link actin to microtubules, as it localizes to the cell cortex in an actin-dependent manner through an interaction with the adherens junction protein, β-catenin46.
Alternatively, structural and regulatory microtubule–actin interactions may mediate specific spatiotemporal regulation of focal contacts with the substrate to guide cell motility (Fig. 2a, panel 2). It has also been shown that during dynamic instability, microtubules specifically target focal contacts, and that targeting frequency is inversely proportional to focal contact lifetime47,48,49. Further evidence indicates that a kinesin microtubule motor may deliver a regulatory factor that promotes focal adhesion disassembly50. Microtubules are probably guided to focal adhesions by structural links to adhesion-associated actin filaments20,49. Indeed, this may be mediated by a similar molecular mechanism as that driving spindle reorientation in yeast, where elegant experiments have shown that Myo2, linked to microtubule plus ends by Kar9 and Bim1, can pull a microtubule along an actin bundle to its specific anchor site in the daughter cell42. However, microtubules are more likely to link to adhesion-associated actin by static crosslinkers and grow along filament bundles.
Microtubule–actin interactions in neuronal growth cone guidance
There is increasing evidence that interactions between actin and microtubules are important for neuronal pathfinding. The region of neurons that mediates pathfinding is the highly motile and actin-rich growth cone at the terminus of the axon. It is well established that actin dynamics are necessary for axon guidance51, although unguided axon growth can occur in the absence of actin assembly52. Conversely, dampening of microtubule dynamics results in highly motile 'wandering' growth cones that can no longer recognize substrate boundaries, suggesting that microtubule–actin interactions are important for organizing directed motility and related signal transduction53.
The cytoskeleton of the growth cone can be organized into central, transition and peripheral domains (Fig. 2b)54. A large bundle of microtubules predominates in the central domain and a sub-population of dynamic microtubule plus ends penetrates into the peripheral domain, where actin bundles comprise the radial array of filopodia and a dense actin meshwork lamellipodium spans between filopodia. The transition zone lies between the central and peripheral domains, and contains contractile actin bundles (actin arcs) orientated perpendicular to filopodia. Similarly to a migrating cell, actin retrograde flow occurs in both lamellipodia and filopodia at the dynamic leading edge of the growth cone.
The relationship between microtubules and actin in the peripheral and central domain has been the subject of several recent studies. Their interdependence was first realized in studies where acute inhibition of actin assembly resulted in rapid microtubule advance into the peripheral domain, showing that F-actin assembly dynamics strongly affects microtubule organization and might inhibit microtubule advance55. Recently, simultaneous imaging of actin and microtubules revealed that microtubules were indeed transported out of the peripheral domain through their linkage to retrograde actin flow21. This explains the previously observed 'barrier' to microtubule advance exerted by the peripheral lamellipodium. These studies also revealed that actin bundles within filopodia function as guides along which microtubules grow towards the leading edge in the direction of growth cone extension (Fig. 3b)21,56,57, a situation reminiscent of microtubule tracking along actin bundles in migrating cells20. Retrograde flow in filopodial actin bundles also moves microtubules rearwards21, indicating that microtubules may be structurally linked to actin. Retrograde flow may also have a role in microtubule turnover, as microtubules that are moved rearward through their attachment to filopodial actin bundles often 'buckle' and break in the transition zone where contractile actin 'arcs' predominate21, similarly to what occurs in the convergence zone of migrating cells20,26. Actin arcs also interact with microtubules and transport them into the central domain. Interestingly, arc-associated microtubules are less dynamic than those associated with filopodia20.
Local perturbation of actin has revealed that microtubule distribution indeed depends on actin bundles. Regional disruption of actin bundles by drug application on one side of the growth cone results in an inability of microtubules to penetrate into that area of the peripheral domain. Consequently, the growth cone turns away from this region and instead grows in the direction containing stabilized filopodia and their associated extended microtubules57. This suggests that the spatial distribution of filopodia dictates the direction of microtubule extension through structural links between microtubules and actin. Microtubule growth, in turn, may promote further lamellipodial protrusion through regulatory pathways, as local promotion of microtubule growth induces local lamellipodial protrusion and growth cone turning58. It is tempting to speculate that this could be caused by local microtubule growth-mediated activation of Rac1 activity, as proposed for migrating cells. Finally, axon branching also depends on microtubule–actin interactions, as depolymerization of either polymer inhibits the number and length of axon branches59.
So, what molecules might mediate the link between microtubules and actin in neuronal growth cones? Twenty years ago, Pollard and colleagues demonstrated that the microtubule-associated proteins MAP2 and Tau could crosslink microtubules to F-actin in vitro60. More recent evidence indicates that phosphorylation may function as a switch to regulate the association of MAP2c with either the microtubule or the actin cytoskeleton61, or possibly the crosslinking of microtubules to actin, although this has yet to be demonstrated. Alternatively, Kakapo/Shortstop, the Drosophila melanogaster homologue of the plakin MACF is required for neuronal axon extension, and this function depends on its actin- and microtubule-binding domains62.
Microtubule–actin interactions in cellular wound healing
Microtubule–actin interactions may be critical to cellular wound healing. Wounds in individual cells are rapidly repaired by intracellular membrane-fusion-dependent assembly of a 'patching membrane'63. In amphibian oocytes and eggs64,65, and various other cell types (C.A.M. and W.M.B., unpublished observations), the patching membrane is rapidly encircled by an array of actin and myosin-2 that closes like a 'purse string' around the wound site. This array is generated by local assembly of actin filaments and myosin-2 around wound borders, and concurrent cortical flow of pre-existing actin filaments towards the wound66. Similarly, microtubules are assembled into a radial array around wounds by local assembly and transport towards the wound23. Multi-spectral imaging of actin and microtubules in live cells showed that microtubules are transported towards wound borders by associating with flowing actin, suggesting structural links between the two polymers23 (Fig. 3c). Microtubule transport results in buckling and breaking of microtubules as they are contracted into the wound edge, creating a zone of microtubule disassembly at the region of high actin contractility near the wound border23 (Fig. 2c). This is similar to the microtubule buckling and breakage that occurs in the convergence and transition zones of migrating cells and growth cones, respectively. Simultaneously, microtubule ends distal from the wound edge polymerize continuously as their shafts flow towards the wound23, similar to the net polymerization of microtubules at the leading edge of migrating cells or growth cones.
The transport of microtubules by structural links to actin flow is reciprocated by microtubule-mediated regulation of actomyosin activity. Disruption of microtubules before wounding impairs the formation of the purse-string array by disrupting the actomyosin array around wound borders67. Conversely, microtubule disruption after the establishment of the actomyosin array accelerates purse-string contraction65. Preliminary work showing localization of GTP-bound RhoA at the wound border, with GTP-bound Rac1 concentrated in a ring distal from the wound border suggests that effects of microtubule perturbations might be mediated by Rho proteins68.
Microtubule–actin interactions in cell division
Some interesting examples of microtubule–actin interactions occur during animal cell division, including spindle positioning and cytokinesis. During mitosis, two MTOCs organize microtubules into two asters that interdigitate into an antiparallel array and interact with kinetochores on replicated sister chromosomes to separate them equally into daughter cells. The majority of actin remains in a cortical network, where it mediates spindle positioning early in mitosis. A small proportion of actin is found among the spindle microtubules69, although the function of actin in the spindle is unclear. Some evidence indicates it may comprise a supramolecular 'spindle matrix,' whereas other evidence suggests it is dispensible. Next, after anaphase chromosome segregation and during cytokinesis, a contractile actomyosin ring assembles to mediate the formation of a furrow in the plasma membrane precisely between the spindle poles to separate the two daughter cells.
Spindles can often be physically linked to specific regions of the cell cortex to ensure that the future division plane is properly placed, especially in asymmetric divisions that are frequent during early development70. This linkage is thought to be mediated by proteins at spindle microtubule plus ends that bind to cortical actin, as now established for the Myo2–Kar9–Bim1 and Cdc42–cytoplasmic dynein pathways in Saccharomyces cerevisiae (above), and is hypothesized to occur during MTOC reorientation in migrating cells39,43. Similarly, in two-cell Caenorhabditis elegans embryos, the cytoplasmic dynein–dynactin complex may capture astral microtubules to orientate the spindle71,72 through interaction with an actin array at a site defined by the previous division plane73. In Drosophila syncytial embryos, spindle anchoring at the cortex depends on armadillo and the Drosophila homologue of APC, both of which may form a complex with α-catenin associated with cortical actin74.
The idea that microtubule–actin interactions are critical during cytokinesis was considered even before microtubules, actin filaments and myosin-2 were characterized. Classic studies showed that spindle disruption before cytokinetic furrow formation prevented cytokinesis, whereas spindle disruption after furrow initiation did not prevent furrow ingression75. However, identifying the molecular mechanism underlying this influence of microtubules on furrow assembly and position is challenging, because microtubules are competent to direct furrowing only during a 1-h window termed the 'C phase' of the cell cycle76. C phase begins after the onset of anaphase76,77 and is characterized by increased cortical contractility. There is also cell-type specificity: in embryos, astral microtubules control furrow position, whereas in smaller cells, microtubules of the spindle midzone are important78. Despite this complexity, it seems that the structural and regulatory interactions between microtubules and actin that operate in other cellular contexts may also contribute to cytokinesis.
Structural interactions between microtubules and actin may mediate furrow positioning. In the simplest scenario, microtubules could directly transport actin and myosin-2. In support of this, in Xenopus egg extracts, actin is transported towards the periphery of asters by binding to microtubules that are moved by cytoplasmic dynein centrifugally from the aster centre17,79. Similarly, analysis of microtubules, actin and myosin-2 in Drosophila embryos suggests that microtubules may transport actomyosin away from MTOCs in a cell cycle-dependent manner80. The centrifugal transport from MTOCs could result in accumulation of actomyosin between two adjacent MTOCs to directly promote assembly of the cytokinetic contractile apparatus81. Candidates for mediating these structural interactions between microtubules and actin include anillin and the septins. Both have microtubule and actin affinity, are localized to the cytokinetic apparatus, and are required for cytokinesis in several systems82,83,84.
Microtubule–actin interactions may also be important for formation of the spindle midzone, a subset of stabilized microtubules that interdigitate between the recently segregated chromosomes after anaphase, and which may contain cytokinesis regulatory signals. For example, disrupting actin through mutation in actin regulatory genes or pharmacological agents causes defects in midzone microtubule organization85,86,87. In addition, multispectral imaging of microtubules and actin suggests that a sub-population of microtubule plus ends attach to specific cortical regions concurrent with cortical actin flow towards the incipient furrow, sweeping the attached microtubules into the midzone regions (J.C. Canman and E.D. Salmon & C.M.W.-S., unpublished observations). However, the molecules that mediate the microtubule/cortex attachment are unknown.
Microtubules may also control actomyosin during cytokinesis by directing the position of Rho family GTPase signalling in a similar manner to that described for cell locomotion: that is, that local microtubule polymerization or high local microtubule density promotes local Rac1 activation15; and local microtubule depolymerization or reduced local microtubule density promotes local RhoA activation14. Specifically, it was proposed that regions flanking the furrow have a high density of astral microtubules that would activate Rac to promote the assembly and subsequent flow of cortical actin filaments into the forming contractile array. In contrast, the furrow region (which is further away from both asters) has a lower density of microtubules and thus high Rho activity, which promotes contraction between the asters81. Consistent with this model is the dependence of cytokinesis on Rho and Rho regulators78, and the genetic interaction between the Rho pathway and pathways that control microtubule disruption during cytokinesis88. In Drosophila, Rho proteins may also mediate structural links between the actin cortex and microtubules through a complex between a Rho GEF, a putative Rac inhibitor and a kinesin, Pavarotti/CHO1 (refs 89, 90).
Alternatively, microtubules could have regulatory interactions with actin that are distinct from those mediated by Rho proteins and may be cell-cycle-specific. For example, 'passenger proteins' such as aurora B kinase may be transported by microtubules from the kinetochores to the spindle midzone and cytokinetic apparatus, where they regulate actomyosin to promote successful furrowing during cytokinesis78,91. This transport could be mediated by the kinesin microtubule motor, CHO1, as it localizes to the midzone and has an actin-binding domain92.
Microtubule–actin interactions in cortical flow
In non-adherent cells, such as oocytes, eggs and embryos, cortical actomyosin contractility is often manifest as cortical flow: the translocation of cortical actin and associated material towards sites of high contractility and/or away from low contractility regions. Cortical flow is powered by asymmetries in contractility within the presumably continuous network of actin and myosin-2 that underlies the plasma membrane, which are spatially controlled by microtubules.
For example, in interphase C. elegans zygotes, the striking actin- and myosin-2-dependent cortical flow away from the future posterior pole is microtubule-dependent. Here, cortical flow controls localization of germ cell determinants and polarity markers through compensatory flow of central cytoplasm towards the future posterior pole. Because the flow is directed away from the large microtubule aster of the sperm93, this suggests that microtubules locally relax the cortex to direct flow away from regions of high microtubule density, as is hypothesized to occur during cytokinesis. Furthermore, in Spd-2 mutants, where sperm aster assembly is disrupted, cortical flow is also perturbed94. Conversely, manipulations that limit sperm aster development but permit growth of microtubules from the female pronucleus at the opposite end of the embyro reverse normal patterns of polarity marker distribution95. Cortical flow can also be induced later in the cell cycle (M phase) by manipulations that displace the spindle such that cortical flow is directed away from the spindle, independent of its location93.
Similarly, cortical flow to the animal pole in Xenopus oocytes also depends on the local level of microtubule polymer96. Displacement of the oocyte nucleus, which functions as a microtubule-organizing centre, results in local flow away from the displaced nucleus97. Together, these results suggest that microtubules direct cortical flow by local inhibition of actomyosin-based contractility, indicating a regulatory interaction between microtubules and actin.
Conserved microtubule/actin mechanochemical activity modules working in varied cellular contexts
One of the most surprising findings to emerge from our 'compare and contrast' analysis is the striking similarity of microtubule–actin interactions in apparently diverse cell systems and morphogenic functions. We propose that microtubule–actin interactions can be distilled into, at the very least, three conserved 'mechanochemical modules' that can be inserted into different cellular contexts and applied to different functions, requiring the establishment and/or maintenance of dynamic cellular asymmetry. By module, we mean small conserved subsets of molecular building blocks that form relatively autonomous, intermediate-scale entities, each with its own intrinsic properties. Microtubule–actin interaction modules may involve either 'structural' or 'regulatory' interactions, or can consist of both. Modules may be put to work singly in simple systems, or multiple modules may cooperate or feed back on one another in more complex cell functions or in bigger cells. We hypothesize that core structural and regulatory molecular components of each module are highly conserved, but that in different cell contexts, modules may have alternative structural or regulatory components to be customized to the specific morphogenic function where the module is put to use. By analogy, the cam is a 'mechanical module' that, depending on its size and shape, can be used to open intake and exhaust valves in a combustion engine, or to push sheets of paper through a photocopy machine; both functions that require the regular periodic application of pressure and/or directed motion for a prescribed length of time.
The most well-characterized microtubule–actin interaction module is what we will call the 'plus end/cortex anchor' (Fig. 4b). Here, plus ends of individual microtubules become bound by stable attachments to sites in the actin cortex defined by Cdc42. This serves as a way of polarizing the centrosome or the mitotic spindle within the cell. As noted, this has been well characterized at the molecular level in S. cerevisiae and discussed in recent reviews39,43,98. We speculate that the same module is at work in positioning centrosomes in migrating cells or insect embryo syncytia, and spindles in asymmetric cell divisions in embryos or polarized epithelia. How highly conserved the members of the yeast 'anchor' module are, and what other molecules participate in this module in higher organisms remain to be determined39.

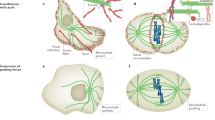
(a) The 'polymerization/contraction treadmill' module may operate in migrating cells, neuronal growth cones and Xenopus oocyte wounds. In this module, a region at the leading edge or distal from the wound border of high Rac1 activity stimulates actin and microtubule polymerization, and is perpetuated by microtubule growth. Microtubules (green) that are crosslinked to actin (red) are moved away from this region by actin flow. As they approach the convergence zone/transition zone or the wound edge, a region of high actin density and high contractility, they buckle and break, generating free minus ends that depolymerize. Microtubule disassembly may activate RhoA to perpetuate actomyosin contraction and stabilization of a subset of microtubules (blue). (b) The 'plus end/cortex anchor' module is well characterized in yeast, where stable attachment of microtubules to the actin cortex is important for positioning the mitotic spindle. Microtubules may attach to the actin cortex and find the bud tip by a complex consisting of the microtubule-end-binding protein Bim1, Kar9 and Myo2, and the spindle may then be positioned in a cytoplasmic dynein-dependent manner. Cytoplasmic dynein linked to the cortex by β-catenin may be important for MTOC orientation during cell migration. (c) The 'actin bundle/microtubule guidance' module may be important for precise positioning of individual microtubules in migrating cells and neuronal growth cones. Here, microtubules may be structurally linked to actin bundles and grow along them.
The second activity module is the 'polymerization/contraction treadmill', which may be used in systems where gradients in polymerization and contractility must be perpetuated (Fig. 4a). This 'treadmill' module comprises both structural and regulatory microtubule–actin interactions. Here, a cellular zone of actin meshwork polymerization and microtubule growth are separated by 5–20 μm from a zone of actomyosin contraction that continually reels in the growing actin meshwork, and with it the attached microtubules. Plus-end microtubule growth at one end of the treadmill offsets the microtubule flow towards the region of contraction, where microtubules are subjected to compressive forces and are buckled, broken and then shorten at their broken minus ends. The 'treadmill' may be maintained by microtubule growth-mediated activation of Rac1 at one end, and broken microtubule shortening-mediated activation of RhoA at the opposite end. Indeed, it was surprising to realize that a cellular wound resembles the leading edge and convergence/transition zone of locomoting cells and neuronal growth cones, respectively (Fig. 3). The treadmill module also most probably helps position the contractile ring during cytokinesis or generate asymmetries in contractility at the cell cortex and thereby power cortical flow. Furthermore, the 'anchor' module and the 'treadmill' module may function cooperatively during cell migration or cytokinesis. Here, the 'anchor' module may initiate a polarization of microtubules towards the leading edge or to cortical regions adjacent to spindle poles. This, in turn, could initiate the activity of the treadmill module to activate locally a self-perpetuating cycle of protrusion and retrograde flow in migrating cells, or polar relaxation and furrow contraction in dividing cells. A challenge for the future is to determine the multiple conserved and variable molecular players in this complex 'polymerization/contraction treadmill' module.
The final putative conserved microtubule–actin interaction module that we define here is the 'actin bundle/microtubule guidance' module, which may be used in situations where single microtubules must be targeted to precise positions (Fig. 4c). Here, microtubules bind to, and grow or move along, bundles of actin filaments. Clearly, this is critical for neuronal pathfinding, where the spatial distribution of filopodia guides the growth of microtubules, which in turn steers growth cone motility. This module is also probably used for microtubule targeting to, and regulation of, focal adhesions in migrating cells. It is also possible that actin in the mitotic spindle could facilitate the targeting of microtubules to kinetochores on chromosomes. This guidance module can be used cooperatively with the 'anchor' module, such as in yeast, where actin cables guide microtubules to cortical anchor sites during cell division42. Microtubule guidance along actin bundles may use either static (probably in neurons or tissue cells) or dynamic motor-mediated (as in yeast) links between microtubules and actin filaments. Here, the critical questions are the mechanism by which actin bundles form and the identity of the structural microtubule–actin crosslinking proteins in various systems.
Finally, we hypothesize that these (and possibly other unknown microtubule–actin interaction modules) may be important in other dynamic morphogenic processes, such as the polarization of T cells towards antigen presenting cells during the immune response99, cellularization in syncytial insect embryos80 and the generation of tubules from epithelial cells during organ development100.
References
Vasiliev, J.M. et al. Effect of colcemid on the locomotory behaviour of fibroblasts. J. Embryol. Exp. Morphol. 24, 625–640 (1970).
Ingber, D.E. Tensegrity II. How structural networks influence cellular information processing networks. J. Cell Sci. 116, 1397–1408 (2003).
Ingber, D.E. Tensegrity I. Cell structure and hierarchical systems biology. J. Cell Sci. 116, 1157–1173 (2003).
Wittmann, T. & Waterman-Storer, C.M. Cell motility: can Rho GTPases and microtubules point the way? J. Cell Sci. 114, 3795–3803 (2001).
Etienne-Manneville, S. & Hall, A. Rho GTPases in cell biology. Nature 420, 629–635 (2002).
Cook, T.A., Nagasaki, T. & Gundersen, G.G. Rho guanosine triphosphatase mediates the selective stabilization of microtubules induced by lysophosphatidic acid. J. Cell Biol. 141, 175–185 (1998).
Ridley, A.J. Rho GTPases and cell migration. J. Cell Sci. 114, 2713–2722 (2001).
Pruyne, D. et al. Role of formins in actin assembly: nucleation and barbed-end association. Science 297, 612–615 (2002).
Sagot, I., Rodal, A.A., Moseley, J., Goode, B.L. & Pellman, D. An actin nucleation mechanism mediated by Bni1 and profilin. Nature Cell Biol. 4, 626–631 (2002).
Palazzo, A.F., Cook, T.A., Alberts, A.S. & Gundersen, G.G. mDia mediates Rho-regulated formation and orientation of stable microtubules. Nature Cell Biol. 3, 723–729 (2001).
Small, J.V., Stradal, T., Vignal, E. & Rottner, K. The lamellipodium: where motility begins. Trends Cell Biol. 12, 112–120 (2002).
Wittmann, T., Bokoch, G.M. & Waterman-Storer, C. Regulation of leading edge microtubule and actin dynamics downstream of Rac1. J. Cell Biol. 161, 845–851 (2003).
Daub, H., Gevaert, K., Vandekerckhove, J., Sobel, A. & Hall, A. Rac/Cdc42 and p65PAK regulate the microtubule-destabilizing protein stathmin through phosphorylation at serine 16. J. Biol.Chem. 276, 1677–1680 (2001).
Ren, X.D., Kiosses, W.B. & Schwartz, M.A. Regulation of the small GTP-binding protein Rho by cell adhesion and the cytoskeleton. EMBO J. 18, 578–585 (1999).
Waterman-Storer, C.M., Worthylake, R.A., Liu, B.P., Burridge, K. & Salmon, E.D. Microtubule growth activates Rac1 to promote lamellipodial protrusion in fibroblasts. Nature Cell Biol. 1, 45–50 (1999).
Krendel, M., Zenke, F.T. & Bokoch, G.M. Nucleotide exchange factor GEF-H1 mediates cross-talk between microtubules and the actin cytoskeleton. Nature Cell Biol. 4, 294–301 (2002).
Waterman-Storer, C. et al. Microtubules remodel actomyosin networks in Xenopus egg extracts via two mechanisms of F-actin transport. J. Cell Biol. 150, 361–376 (2000).
Leung, C.L., Green, K.J. & Liem, R.K. Plakins: a family of versatile cytolinker proteins. Trends Cell Biol. 12, 37–45 (2002).
Wiche, G. Role of plectin in cytoskeleton organization and dynamics. J. Cell Sci. 111, 2477–2486 (1998).
Salmon, W.C., Adams, M.C., & Waterman-Storer, C.M. Dual-wavelength fluorescent speckle microscopy reveals coupling of microtubule and actin movements in migrating cells. J. Cell Biol. 158, 31–37 (2002).
Schaefer, A.W., Kabir, N., & Forscher, P. Filopodia and actin arcs guide the assembly and transport of two populations of microtubules with unique dynamic parameters in neuronal growth cones. J. Cell Biol. 158, 139–152 (2002).
Mandato, C.A., Weber, K.L., Zandy, A.J., Keating, T.J., & Bement, W.M. Xenopus egg extracts as a model system for analysis of microtubule, actin filament, and intermediate filament interactions. Methods Mol. Biol. 161, 229–239 (2001).
Mandato, C.A. & Bement, W.M. Actomyosin transports microtubules while microtubules control actomyosin recruitment during Xenopus oocyte wound healing. Curr. Biol. (in the press).
Cramer, L.P. Organization and polarity of actin filament networks in cells: implications for the mechanism of myosin-based cell motility. Biochem. Soc. Symp. 65, 173–205 (1999).
Cramer, L.P., Siebert, M. & Mitchison, T.J. Identification of novel graded polarity actin filament bundles in locomoting heart fibroblasts: implications for the generation of motile force. J. Cell Biol. 136, 1287–1305 (1997).
Gupton, S.L., Salmon, W.C. & Waterman-Storer, C. Converging populations of f-actin promote breakage of associated microtubules to spatially regulate microtubule turnover in migrating cells. Curr. Biol. 12, 1891–1899 (2002).
Webb, D.J., Parsons, J.T. & Horwitz, A.F. Adhesion assembly, disassembly and turnover in migrating cells — over and over and over again. Nature Cell Biol. 4, E97–E100 (2002).
Waterman-Storer, C.M. & Salmon, E.D. Actomyosin-based retrograde flow of microtubules in the lamella of migrating epithelial cells influences microtubule dynamic instability and turnover and is associated with microtubule breakage and treadmilling. J. Cell Biol. 139, 417–434 (1997).
Yvon, A.M. & Wadsworth, P. Region-specific microtubule transport in motile cells. J. Cell Biol. 151, 1003–1012 (2000).
Mikhailov, A.V. & Gundersen, G.G. Centripetal transport of microtubules in motile cells. Cell Motil. Cytoskeleton 32, 173–186 (1995).
Wadsworth, P. Regional regulation of microtubule dynamics in polarized, motile cells. Cell Motil. Cytoskeleton 42, 48–59 (1999).
Kraynov, V.S. et al. Localized Rac activation dynamics visualized in living cells. Science 290, 333–337 (2000).
Itoh, R.E. et al. Activation of rac and cdc42 video imaged by fluorescent resonance energy transfer-based single-molecule probes in the membrane of living cells. Mol. Cell Biol. 22, 6582–6591 (2002).
Barth, A.I. & Nelson, W.J. What can humans learn from flies about adenomatous polyposis coli? Bioessays 24, 771–774 (2002).
Nathke, M. et al. The adenomatous polyposis coli tumor suppressor protein localizes to plasma membrane sites involved in active cell migration. J. Cell Biol. 134, 165–179 (1996).
Kawasaki, Y. et al. Asef, a link between the tumor suppressor APC and G-protein signaling. Science 289, 1194–1197 (2000).
Kawasaki, Y., Sato, R. & Akiyama, T. Mutated APC and Asef are involved in the migration of colorectal tumour cells. Nature Cell Biol. 5, 211–215 (2003).
Fukata, M. et al. Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 109, 873–885 (2002).
Gundersen, G.G. Evolutionary conservation of microtubule-capture mechanisms. Nature Rev. Mol. Cell Biol. 3, 296–304 (2002).
Palazzo, A.F. et al. Cdc42, dynein, and dynactin regulate MTOC reorientation independent of Rho-regulated microtubule stabilization. Curr. Biol. 11, 1536–1541 (2001).
Etienne-Manneville, S. & Hall, A. Integrin-mediated activation of Cdc42 controls cell polarity in migrating astrocytes through PKCzeta. Cell 106, 489–498 (2001).
Hwang, E., Kusch, J., Barral, Y. & Huffaker, T.C. Spindle orientation in Saccharomyces cerevisiae depends on the transport of microtubule ends along polarized actin cables. J. Cell Biol. 161, 483–488 (2003).
Yarm, F., Sagot, I. & Pellman, D. The social life of actin and microtubules: interaction versus cooperation. Curr. Opin. Microbiol. 4, 696–702 (2001).
Barth, A.I., Siemers, K.A. & Nelson, W.J. Dissecting interactions between EB1, microtubules and APC in cortical clusters at the plasma membrane. J. Cell Sci. 115, 1583–1590 (2002).
Mimori-Kiyosue, Y., Shiina, N. & Tsukita, S. Adenomatous polyposis coli (APC) protein moves along microtubules and concentrates at their growing ends in epithelial cells. J. Cell Biol. 148, 505–518 (2000).
Ligon, L.A., Karki, S., Tokito, M. & Holzbaur, E.L. Dynein binds to beta-catenin and may tether microtubules at adherens junctions. Nature Cell Biol. 3, 913–917 (2001).
Kaverina, I., Rottner, K. & Small, J.V. Targeting, capture, and stabilization of microtubules at early focal adhesions. J. Cell Biol. 142, 181–190 (1998).
Kaverina, I., Krylyshkina, O. & Small, J.V. Microtubule targeting of substrate contacts promotes their relaxation and dissociation. J. Cell Biol. 146, 1033–1044 (1999).
Krylyshkina, O. et al. Nanometer targeting of microtubules to focal adhesions. J. Cell Biol. 161, 853–859 (2003).
Krylyshkina, O. et al. Modulation of substrate adhesion dynamics via microtubule targeting requires kinesin-1. J. Cell Biol. 156, 349–359 (2002).
Bentley, D. & Toroian-Raymond, A. Disoriented pathfinding by pioneer neuron growth cones deprived of filopodia by cytochalasin treatment. Nature 323, 712–715 (1986).
Marsh, L. & Letourneau, P.C. Growth of neurites without filopodial or lamellipodial activity in the presence of cytochalasin B. J. Cell Biol. 99, 2041–2047 (1984).
Tanaka, E., Ho, T. & Kirschner, M.W. The role of microtubule dynamics in growth cone motility and axonal growth. J. Cell Biol. 128, 139–155 (1995).
Suter, D.M. & Forscher, P. An emerging link between cytoskeletal dynamics and cell adhesion molecules in growth cone guidance. Curr. Opin. Neurobiol. 8, 106–116 (1998).
Forscher, P. & Smith, S.J. Actions of cytochalasins on the organization of actin filaments and microtubules in a neuronal growth cone. J. Cell Biol. 107, 1505–1516 (1988).
Kabir, N., Schaefer, A.W., Nakhost, A., Sossin, W.S. & Forscher, P. Protein kinase C activation promotes microtubule advance in neuronal growth cones by increasing average microtubule growth lifetimes. J. Cell Biol. 152, 1033–1044 (2001).
Zhou, F.Q., Waterman-Storer, C.M. & Cohan, C.S. Focal loss of actin bundles causes microtubule redistribution and growth cone turning. J. Cell Biol. 157, 839–849 (2002).
Buck, K.B. & Zheng, J.Q. Growth cone turning induced by direct local modification of microtubule dynamics. J. Neurosci. 22, 9358–9367 (2002).
Dent, E.W. & Kalil, K. Axon branching requires interactions between dynamic microtubules and actin filaments. J. Neurosci. 21, 9757–9769 (2001).
Griffith, L.M. & Pollard, T.D. The interaction of actin filaments with microtubules and microtubule- associated proteins. J. Biol.Chem. 257, 9143–9151 (1982).
Ozer, R.S. & Halpain, S. Phosphorylation-dependent localization of microtubule-associated protein MAP2c to the actin cytoskeleton. Mol. Biol. Cell 11, 3573–3587 (2000).
Lee, S. & Kolodziej, P.A. Short Stop provides an essential link between F-actin and microtubules during axon extension. Development 129, 1195–1204 (2002).
McNeil, P.L. & Terasaki, M. Coping with the inevitable: how cells repair a torn surface membrane. Nature Cell Biol. 3, E124–E129 (2001).
Merriam, R.W. & Christensen, K. A contractile ring-like mechanism in wound healing and soluble factors affecting structural stability in the cortex of Xenopus eggs and oocytes. J. Embryol. Exp. Morphol. 75, 11–20 (1983).
Bement, W.M., Mandato, C.A. & Kirsch, M.N. Wound-induced assembly and closure of an actomyosin purse string in Xenopus oocytes. Curr. Biol. 9, 579–587 (1999).
Mandato, C.A. & Bement, W.M. Contraction and polymerization cooperate to assemble and close actomyosin rings around Xenopus oocyte wounds. J. Cell Biol. 154, 785–797 (2001).
Mandato, C.A. & Bement, W.M. Transport and polymerization interactions between microtubules and F-actin during Xenopus oocyte wound healing. Mol. Biol. Cell 13, 333a (2002).
Benink, H.A. & Bement, W.M. Concentric rings of rho, rac and Cdc42 activity encircle oocyte wounds. Mol. Biol. Cell 13, 36a (2002).
Silverman-Gavrila, R.V. & Forer, A. Evidence that actin and myosin are involved in the poleward flux of tubulin in metaphase kinetochore microtubules of crane-fly spermatocytes. J. Cell Sci. 113, 597–609 (2000).
Lutz, D.A., Hamaguchi, Y. & Inoue, S. Micromanipulation studies of the asymmetric positioning of the maturation spindle in Chaetopterus sp. oocytes: I. Anchorage of the spindle to the cortex and migration of a displaced spindle. Cell Motil. Cytoskeleton 11, 83–96 (1988).
Skop, A.R. & White, J.G. The dynactin complex is required for cleavage plane specification in early Caenorhabditis elegans embryos. Curr. Biol. 8, 1110–1116 (1998).
Gonczy, P., Pichler, S., Kirkham, M. & Hyman, A.A. Cytoplasmic dynein is required for distinct aspects of MTOC positioning, including centrosome separation, in the one cell stage Caenorhabditis elegans embryo. J. Cell Biol. 147, 135–150 (1999).
Hyman, A.A. & White, J.G. Determination of cell division axes in the early embryogenesis of Caenorhabditis elegans. J. Cell Biol. 105, 2123–2135 (1987).
McCartney, B.M. et al. Drosophila APC2 and Armadillo participate in tethering mitotic spindles to cortical actin. Nature Cell Biol. 3, 933–938 (2001).
Hiramoto, Y. Cell division without mitotic apparatus in sea urchin eggs. Exp. Cell Res. 11, 630–636 (1956).
Canman, J.C., Hoffman, D.B. & Salmon, E.D. The role of pre- and post-anaphase microtubules in the cytokinesis phase of the cell cycle. Curr. Biol. 10, 611–614 (2000).
Shuster, C.B. & Burgess, D.R. Transitions regulating the timing of cytokinesis in embryonic cells. Curr. Biol. 12, 854–858 (2002).
Glotzer, M. Animal cell cytokinesis. Annu. Rev. Cell Dev. Biol. 17, 351–386 (2001).
Sider, J.R. et al. Direct observation of microtubule–F-actin interaction in cell free lysates. J. Cell Sci. 112, 1947–1956 (1999).
Foe, V.E., Field, C.M. & Odell, G.M. Microtubules and mitotic cycle phase modulate spatiotemporal distributions of F-actin and myosin II in Drosophila syncytial blastoderm embryos. Development 127, 1767–1787 (2000).
Mandato, C.A., Benink, H.A. & Bement, W.M. Microtubule-actomyosin interactions in cortical flow and cytokinesis. Cell Motil. Cytoskeleton 45, 87–92 (2000).
Sisson, J.C., Field, C., Ventura, R., Royou, A. & Sullivan, W. Lava lamp, a novel peripheral Golgi protein, is required for Drosophila melanogaster cellularization. J. Cell Biol. 151, 905–918 (2000).
Oegema, K., Savoian, M.S., Mitchison, T.J. & Field, C.M. Functional analysis of a human homologue of the Drosophila actin binding protein anillin suggests a role in cytokinesis. J. Cell Biol. 150, 539–552 (2000).
Surka, M.C., Tsang, C.W. & Trimble, W.S. The mammalian septin MSF localizes with microtubules and is required for completion of cytokinesis. Mol. Biol. Cell 13, 3532–3545 (2002).
Giansanti, M.G. et al. Cooperative interactions between the central spindle and the contractile ring during Drosophila cytokinesis. Genes Dev. 12, 396–410 (1998).
Cimini, D., Fioravanti, D., Tanzarella, C. & Degrassi, F. Simultaneous inhibition of contractile ring and central spindle formation in mammalian cells treated with cytochalasin B. Chromosoma 107, 479–485 (1998).
Somma, M.P., Fasulo, B., Cenci, G., Cundari, E. & Gatti, M. Molecular dissection of cytokinesis by RNA interference in Drosophila cultured cells. Mol. Biol. Cell 13, 2448–2460 (2002).
Dechant, R. & Glotzer, M. Centrosome separation and central spindle assembly act in redundant pathways that regulate microtubule density and trigger cleavage furrow formation. Dev. Cell 4, 333–344 (2003).
Somers, W.G. & Saint, R. A RhoGEF and Rho Family GTPase-Activating Protein Complex Links the Contractile Ring to Cortical Microtubules at the Onset of Cytokinesis. Dev. Cell 4, 29–39 (2003).
Adams, R.R., Tavares, A.A., Salzberg, A., Bellen, H.J. & Glover, D.M. pavarotti encodes a kinesin-like protein required to organize the central spindle and contractile ring for cytokinesis. Genes Dev. 12, 1483–1494 (1998).
Murata-Hori, M. & Wang, Y.L. Both midzone and astral microtubules are involved in the delivery of cytokinesis signals: insights from the mobility of aurora B. J. Cell Biol. 159, 45–53 (2002).
Kuriyama, R., Gustus, C., Terada, Y., Uetake, Y. & Matuliene, J. CHO1, a mammalian kinesin-like protein, interacts with F-actin and is involved in the terminal phase of cytokinesis. J. Cell Biol. 156, 783–790 (2002).
Hird, S.N. & White, J.G. Cortical and cytoplasmic flow polarity in early embryonic cells of Caenorhabditis elegans. J. Cell Biol. 121, 1343–1355 (1993).
O'Connell, K.F., Maxwell, K.N. & White, J.G. The spd-2 gene is required for polarization of the anteroposterior axis and formation of the sperm asters in the Caenorhabditis elegans zygote. Dev. Biol. 222, 55–70 (2000).
Wallenfang, M.R. & Seydoux, G. Polarization of the anterior-posterior axis of C. elegans is a microtubule-directed process. Nature 408, 89–92 (2000).
Canman, J.C. & Bement, W.M. Microtubules suppress actomyosin-based cortical flow in Xenopus oocytes. J. Cell Sci. 110, 1907–1917 (1997).
Benink, H.A., Mandato, C.A. & Bement, W.M. Analysis of cortical flow models in vivo. Mol. Biol. Cell 11, 2553–2563 (2000).
Dujardin, D.L. & Vallee, R.B. Dynein at the cortex. Curr. Opin. Cell Biol. 14, 44–49 (2002).
Stowers, L., Yelon, D., Berg, L.J. & Chant, J. Regulation of the polarization of T cells toward antigen-presenting cells by Ras-related GTPase CDC42. Proc. Natl Acad. Sci. USA 92, 5027–5031 (1995).
O'Brien, L.E., Zegers, M.M. & Mostov, K.E. Opinion: Building epithelial architecture: insights from three- dimensional culture models. Nature Rev. Mol. Cell Biol. 3, 531–537 (2002).
Goode, B.L. et al. Coronin promotes the rapid assembly and cross-linking of actin filaments and may link the actin and microtubule cytoskeletons in yeast. J. Cell Biol. 144, 83–98 (1999).
Bashour, A.M., Fullerton, A.T., Hart, M.J. & Bloom, G.S. IQGAP1, a Rac- and Cdc42-binding protein, directly binds and cross- links microfilaments. J. Cell Biol. 137, 1555–1566 (1997).
Mateer, S.C. et al. The mechanism for regulation of the F-actin binding activity of IQGAP1 by calcium/calmodulin. J. Biol. Chem. 277, 12324–12333 (2002).
Ruiz-Velasco, R., Lanning, C.C. & Williams, C.L. The activation of Rac1 by M3 muscarinic acetylcholine receptors involves the translocation of Rac1 and IQGAP1 to cell junctions and changes in the composition of protein complexes containing Rac1, IQGAP1, and actin. J. Biol. Chem. 277, 33081–33091 (2002).
Cunningham, C.C. et al. Microtubule-associated protein 2c reorganizes both microtubules and microfilaments into distinct cytological structures in an actin-binding protein-280-deficient melanoma cell line. J. Cell Biol. 136, 845–857 (1997).
Gonzalez, M., Cambiazo, V. & Maccioni, R.B. The interaction of Mip-90 with microtubules and actin filaments in human fibroblasts. Exp. Cell Res. 239, 243–253 (1998).
Huang, J.D. et al. Direct interaction of microtubule- and actin-based transport motors. Nature 397, 267–270 (1999).
Lantz, V.A. & Miller, K.G. A class VI unconventional myosin is associated with a homologue of a microtubule-binding protein, cytoplasmic linker protein-170, in neurons and at the posterior pole of Drosophila embryos. J. Cell Biol. 140, 897–910 (1998).
Hicks, J.L., Deng, W.M., Rogat, A.D., Miller, K.G. & Bownes, M. Class VI unconventional myosin is required for spermatogenesis in Drosophila. Mol. Biol. Cell 10, 4341–4353 (1999).
Goriounov, D., Leung, C.L. & Liem, R.K. Protein products of human Gas2-related genes on chromosomes 17 and 22 (hGAR17 and hGAR22) associate with both microfilaments and microtubules. J. Cell Sci. 116, 1045–1058 (2003).
Acknowledgements
We thank J. Canman, G. von Dassow and members of the Waterman-Storer lab for helpful suggestions. C.W.S. is supported by National Institutes of Health (NIH) grant GM61804-03, W.M.B by NIH grant GM52932-04A1 and National Science Foundation (NSF) grant MCB#9630860, P.F. by NIH grant RO1-NS28695, A.W.S. by NIH and O.C.R. by NSF postdoctoral fellowships, respectively.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rodriguez, O., Schaefer, A., Mandato, C. et al. Conserved microtubule–actin interactions in cell movement and morphogenesis. Nat Cell Biol 5, 599–609 (2003). https://doi.org/10.1038/ncb0703-599
Issue Date:
DOI: https://doi.org/10.1038/ncb0703-599
This article is cited by
-
The Critical Role of the Shroom Family Proteins in Morphogenesis, Organogenesis and Disease
Phenomics (2024)
-
Hax1 regulate focal adhesion dynamics through IQGAP1
Cell Communication and Signaling (2023)
-
Biomechanical, biophysical and biochemical modulators of cytoskeletal remodelling and emergent stem cell lineage commitment
Communications Biology (2023)
-
Silk-derived peptide nanospirals assembled by self-propelled worm-like filaments
Nano Research (2023)
-
Modeling axonal regeneration by changing cytoskeletal dynamics in stem cell-derived motor nerve organoids
Scientific Reports (2022)
