
Abstract
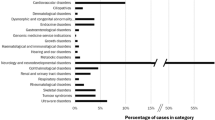
To assess factors influencing the success of whole-genome sequencing for mainstream clinical diagnosis, we sequenced 217 individuals from 156 independent cases or families across a broad spectrum of disorders in whom previous screening had identified no pathogenic variants. We quantified the number of candidate variants identified using different strategies for variant calling, filtering, annotation and prioritization. We found that jointly calling variants across samples, filtering against both local and external databases, deploying multiple annotation tools and using familial transmission above biological plausibility contributed to accuracy. Overall, we identified disease-causing variants in 21% of cases, with the proportion increasing to 34% (23/68) for mendelian disorders and 57% (8/14) in family trios. We also discovered 32 potentially clinically actionable variants in 18 genes unrelated to the referral disorder, although only 4 were ultimately considered reportable. Our results demonstrate the value of genome sequencing for routine clinical diagnosis but also highlight many outstanding challenges.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Accession codes
Primary accessions
European Nucleotide Archive
Referenced accessions
NCBI Reference Sequence
References
Need, A.C. et al. Clinical application of exome sequencing in undiagnosed genetic conditions. J. Med. Genet. 49, 353–361 (2012).
Bamshad, M.J. et al. Exome sequencing as a tool for Mendelian disease gene discovery. Nat. Rev. Genet. 12, 745–755 (2011).
Yang, Y. et al. Clinical whole-exome sequencing for the diagnosis of mendelian disorders. N. Engl. J. Med. 369, 1502–1511 (2013).
Gonzaga-Jauregui, C., Lupski, J.R. & Gibbs, R.A. Human genome sequencing in health and disease. Annu. Rev. Med. 63, 35–61 (2012).
Dixon-Salazar, T.J. et al. Exome sequencing can improve diagnosis and alter patient management. Sci. Transl. Med. 4, 138ra78 (2012).
1000 Genomes Project Consortium. An integrated map of genetic variation from 1,092 human genomes. Nature 491, 56–65 (2012).
Tennessen, J.A. et al. Evolution and functional impact of rare coding variation from deep sequencing of human exomes. Science 337, 64–69 (2012).
Beaulieu, C.L. et al. FORGE Canada Consortium: outcomes of a 2-year national rare-disease gene-discovery project. Am. J. Hum. Genet. 94, 809–817 (2014).
Biesecker, L.G. & Green, R.C. Diagnostic clinical genome and exome sequencing. N. Engl. J. Med. 370, 2418–2425 (2014).
Saunders, C.J. et al. Rapid whole-genome sequencing for genetic disease diagnosis in neonatal intensive care units. Sci. Transl. Med. 4, 154ra135 (2012).
Gilissen, C. et al. Genome sequencing identifies major causes of severe intellectual disability. Nature 511, 344–347 (2014).
Jacob, H.J. et al. Genomics in clinical practice: lessons from the front lines. Sci Transl. Med. 5, 194cm5 (2013).
Cazier, J.B. et al. Whole-genome sequencing of bladder cancers reveals somatic CDKN1A mutations and clinicopathological associations with mutation burden. Nat. Commun. 5, 3756 (2014).
Babbs, C. et al. Homozygous mutations in a predicted endonuclease are a novel cause of congenital dyserythropoietic anemia type I. Haematologica 98, 1383–1387 (2013).
Martin, H.C. et al. Clinical whole-genome sequencing in severe early-onset epilepsy reveals new genes and improves molecular diagnosis. Hum. Mol. Genet. 23, 3200–3211 (2014).
Sharma, V.P. et al. Mutations in TCF12, encoding a basic helix-loop-helix partner of TWIST1, are a frequent cause of coronal craniosynostosis. Nat. Genet. 45, 304–307 (2013).
Cossins, J. et al. Congenital myasthenic syndromes due to mutations in ALG2 and ALG14. Brain 136, 944–956 (2013).
Lise, S. et al. Recessive mutations in SPTBN2 implicate β-III spectrin in both cognitive and motor development. PLoS Genet. 8, e1003074 (2012).
Palles, C. et al. Germline mutations affecting the proofreading domains of POLE and POLD1 predispose to colorectal adenomas and carcinomas. Nat. Genet. 45, 136–144 (2013).
McCarthy, D.J. et al. Choice of transcripts and software has a large effect on variant annotation. Genome Med. 6, 26 (2014).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 38, e164 (2010).
McLaren, W. et al. Deriving the consequences of genomic variants with the Ensembl API and SNP Effect Predictor. Bioinformatics 26, 2069–2070 (2010).
Nelson, M.R. et al. An abundance of rare functional variants in 202 drug target genes sequenced in 14,002 people. Science 337, 100–104 (2012).
Stenson, P.D. et al. The Human Gene Mutation Database: 2008 update. Genome Med. 1, 13 (2009).
Pagel, P. et al. The MIPS mammalian protein-protein interaction database. Bioinformatics 21, 832–834 (2005).
de Ligt, J. et al. Diagnostic exome sequencing in persons with severe intellectual disability. N. Engl. J. Med. 367, 1921–1929 (2012).
Swaminathan, G. & Tsygankov, A.Y. The Cbl family proteins: ring leaders in regulation of cell signaling. J. Cell. Physiol. 209, 21–43 (2006).
Denayer, E. & Legius, E. What's new in the neuro-cardio-facial-cutaneous syndromes? Eur. J. Pediatr. 166, 1091–1098 (2007).
Martinelli, S. et al. Heterozygous germline mutations in the CBL tumor-suppressor gene cause a Noonan syndrome-like phenotype. Am. J. Hum. Genet. 87, 250–257 (2010).
Niemeyer, C.M. et al. Germline CBL mutations cause developmental abnormalities and predispose to juvenile myelomonocytic leukemia. Nat. Genet. 42, 794–800 (2010).
Pérez, B. et al. Germline mutations of the CBL gene define a new genetic syndrome with predisposition to juvenile myelomonocytic leukaemia. J. Med. Genet. 47, 686–691 (2010).
Nava, C. et al. Analysis of the chromosome X exome in patients with autism spectrum disorders identified novel candidate genes, including TMLHE. Transl. Psychiatry 2, e179 (2012).
Isrie, M. et al. HUWE1 mutation explains phenotypic severity in a case of familial idiopathic intellectual disability. Eur. J. Med. Genet. 56, 379–382 (2013).
Froyen, G. et al. Submicroscopic duplications of the hydroxysteroid dehydrogenase HSD17B10 and the E3 ubiquitin ligase HUWE1 are associated with mental retardation. Am. J. Hum. Genet. 82, 432–443 (2008).
McMullin, M.F. The classification and diagnosis of erythrocytosis. Int. J. Lab. Hematol. 30, 447–459 (2008).
Jelkmann, W. Regulation of erythropoietin production. J. Physiol. (Lond.) 589, 1251–1258 (2011).
Bowl, M.R. et al. An interstitial deletion-insertion involving chromosomes 2p25.3 and Xq27.1, near SOX3, causes X-linked recessive hypoparathyroidism. J. Clin. Invest. 115, 2822–2831 (2005).
Zajac, J.D. & Danks, J.A. The development of the parathyroid gland: from fish to human. Curr. Opin. Nephrol. Hypertens. 17, 353–356 (2008).
Green, R.C. et al. ACMG recommendations for reporting of incidental findings in clinical exome and genome sequencing. Genet. Med. 15, 565–574 (2013).
MacArthur, D.G. et al. Guidelines for investigating causality of sequence variants in human disease. Nature 508, 469–476 (2014).
Metcalfe, K. et al. Family history of cancer and cancer risks in women with BRCA1 or BRCA2 mutations. J. Natl. Cancer Inst. 102, 1874–1878 (2010).
Zuk, O. et al. Searching for missing heritability: designing rare variant association studies. Proc. Natl. Acad. Sci. USA 111, E455–E464 (2014).
Moutsianas, L. et al. The power of gene-based rare variant methods to detect disease-associated variation and test hypotheses about complex disease. PLoS Genet. 11, e1005165 (2015).
Kapplinger, J.D. et al. Distinguishing arrhythmogenic right ventricular cardiomyopathy/dysplasia-associated mutations from background genetic noise. J. Am. Coll. Cardiol. 57, 2317–2327 (2011).
Castéra, L. et al. Next-generation sequencing for the diagnosis of hereditary breast and ovarian cancer using genomic capture targeting multiple candidate genes. Eur. J. Hum. Genet. 22, 1305–1313 (2014).
Chong, H.K. et al. The validation and clinical implementation of BRCAplus: a comprehensive high-risk breast cancer diagnostic assay. PLoS ONE 9, e97408 (2014).
Borg, A. et al. Characterization of BRCA1 and BRCA2 deleterious mutations and variants of unknown clinical significance in unilateral and bilateral breast cancer: the WECARE study. Hum. Mutat. 31, E1200–E1240 (2010).
Rebbeck, T.R. et al. Bilateral prophylactic mastectomy reduces breast cancer risk in BRCA1 and BRCA2 mutation carriers: the PROSE Study Group. J. Clin. Oncol. 22, 1055–1062 (2004).
Håkansson, S. et al. Moderate frequency of BRCA1 and BRCA2 germ-line mutations in Scandinavian familial breast cancer. Am. J. Hum. Genet. 60, 1068–1078 (1997).
Landrum, M.J. et al. ClinVar: public archive of relationships among sequence variation and human phenotype. Nucleic Acids Res. 42, D980–D985 (2014).
Caputo, S. et al. Description and analysis of genetic variants in French hereditary breast and ovarian cancer families recorded in the UMD-BRCA1/BRCA2 databases. Nucleic Acids Res. 40, D992–D1002 (2012).
Brohet, R.M. et al. Breast and ovarian cancer risks in a large series of clinically ascertained families with a high proportion of BRCA1 and BRCA2 Dutch founder mutations. J. Med. Genet. 51, 98–107 (2014).
Moss, A.J. et al. Clinical aspects of type-1 long-QT syndrome by location, coding type, and biophysical function of mutations involving the KCNQ1 gene. Circulation 115, 2481–2489 (2007).
Choi, G. et al. Spectrum and frequency of cardiac channel defects in swimming-triggered arrhythmia syndromes. Circulation 110, 2119–2124 (2004).
Kapplinger, J.D. et al. Spectrum and prevalence of mutations from the first 2,500 consecutive unrelated patients referred for the FAMILION long QT syndrome genetic test. Heart Rhythm 6, 1297–1303 (2009).
Crotti, L. et al. Long QT syndrome–associated mutations in intrauterine fetal death. J. Am. Med. Assoc. 309, 1473–1482 (2013).
Li, Y. et al. Intracellular ATP binding is required to activate the slowly activating K+ channel IKs . Proc. Natl. Acad. Sci. USA 110, 18922–18927 (2013).
Vukcevic, M. et al. Functional properties of RYR1 mutations identified in Swedish patients with malignant hyperthermia and central core disease. Anesth. Analg. 111, 185–190 (2010).
Lamble, S. et al. Improved workflows for high throughput library preparation using the transposome-based Nextera system. BMC Biotechnol. 13, 104 (2013).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Lunter, G. & Goodson, M. Stampy: a statistical algorithm for sensitive and fast mapping of Illumina sequence reads. Genome Res. 21, 936–939 (2011).
Rimmer, A. et al. Integrating mapping-, assembly- and haplotype-based approaches for calling variants in clinical sequencing applications. Nat. Genet. 46, 912–918 (2014).
Pagnamenta, A.T. et al. Exome sequencing can detect pathogenic mosaic mutations present at low allele frequencies. J. Hum. Genet. 57, 70–72 (2012).
Ruark, E. et al. Mosaic PPM1D mutations are associated with predisposition to breast and ovarian cancer. Nature 493, 406–410 (2013).
Thorvaldsdóttir, H., Robinson, J.T. & Mesirov, J.P. Integrative Genomics Viewer (IGV): high-performance genomics data visualization and exploration. Brief. Bioinform. 14, 178–192 (2013).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 38, e164 (2010).
Yau, C. OncoSNP-SEQ: a statistical approach for the identification of somatic copy number alterations from next-generation sequencing of cancer genomes. Bioinformatics 29, 2482–2484 (2013).
Plagnol, V. et al. A robust model for read count data in exome sequencing experiments and implications for copy number variant calling. Bioinformatics 28, 2747–2754 (2012).
McQuillan, R. et al. Runs of homozygosity in European populations. Am. J. Hum. Genet. 83, 359–372 (2008).
Colella, S. et al. QuantiSNP: an Objective Bayes Hidden-Markov Model to detect and accurately map copy number variation using SNP genotyping data. Nucleic Acids Res. 35, 2013–2025 (2007).
Abecasis, G.R., Cherny, S.S., Cookson, W.O. & Cardon, L.R. Merlin—rapid analysis of dense genetic maps using sparse gene flow trees. Nat. Genet. 30, 97–101 (2002).
Quinlan, A.R. & Hall, I.M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841–842 (2010).
Diez-Roux, G. et al. A high-resolution anatomical atlas of the transcriptome in the mouse embryo. PLoS Biol. 9, e1000582 (2011).
Acknowledgements
We thank the patients and their families who consented to these studies and the High-Throughput Genomics Group at the Wellcome Trust Centre for Human Genetics for the generation of the sequencing data. Additionally, we are grateful to F. Harrington, C. Mignion, V. Sharma, I. Taylor and I. Westbury for assistance with molecular genetic analysis and the staff of the Oxford University Hospitals Regional Genetics and Immunology Laboratories for the DNA preparation for some of the samples.
This work was funded by a Wellcome Trust Core Award (090532/Z/09/Z) and a Medical Research Council Hub grant (G0900747 91070) to P.D., the NIHR Biomedical Research Centre Oxford, the UK Department of Health's NIHR Biomedical Research Centres funding scheme and Illumina. Additional support is acknowledged from the Biotechnology and Biological Science Research Council (BBSRC) (BB/I02593X/1) to G.L. and G.M.; Wellcome Trust grants 093329, 091182 and 102731 to A.O.M.W. and 100308 to L.F.; the Newlife Foundation for Disabled Children (10-11/04) to A.O.M.W.; AtaxiaUK to A.H.N.; the Haemochromatosis Society to K.R.; European Research Council (FP7/2007-2013) grant agreements 281824 to J.C.K. and 305608 to O.D.; the Jeffrey Modell Foundation NYC and Baxter Healthcare to S.Y.P. and H. Chapel; Action de Recherche Concertée (ARC10/15-029, Communauté Française de Belgique) to O.D.; Fonds de la Recherche Scientifique (FNRS), Fonds de la Recherche Scientifique Médicale (FRSM) and Inter-University Attraction Pole (IUAP; Belgium federal government) to O.D.; the Swiss National Centre of Competence in Research Kidney Control of Homeostasis Program to O.D.; the Gebert Rüf Stiftung (project GRS-038/12) to O.D.; Swiss National Science Foundation grant 310030-146490 to O.D.; the Shriners Hospitals for Children (grant 15958) to M.P.W.; and UK Medical Research Council grants G9825289 and G1000467 to R.V.T., L009609 to A.R.A., G1000801 to D.H. and MC_UC_12010/3 to L.F. The views expressed in this publication are those of the authors and not necessarily those of the UK Department of Health.
Author information
Authors and Affiliations
Contributions
P.D. and G.M. jointly supervised and oversaw the WGS500 project. C. Babbs, D. Beeson, P.B., E.B., H. Chapel, R.C., J.F., L.F., D.H., A.H., F.K., U.K., J.C.K., A.H.N., S.Y.P., C.P., F.P., P.J.R., P.A.R., K.R., A. Schuh, A. Simmons, R.V.T., I.T., H.H.U., P.V., H.W. and A.O.M.W. were principal investigators on individual projects. V.J.B., K.B., C.D., O.D., R.D.G., J.K., C.L., M.A.N., N. Petousi, S.E.P., S.R.F.T., T.V. and M.P.W. were lead investigators on individual projects. H. Cario, M.F.M., C. Bento, K.D., O.D., R.D.G., D.J., C.L., D.N., E.O., A.B.O., M.P., A. Russo, E. Silverman, P.S.S., E. Sweeney, S.A.W. and M.P.W. contributed clinical samples and clinical data. C.A., M.A., A. Green, S.H., Z.K., S. Lamble, L.L., P.P., G.P.-E., A.T. and L.W. prepared libraries and generated whole-genome sequences, led by D. Buck (High-Throughput Genomics Group, Oxford) and D. Bentley (Illumina Cambridge). J. Becq, J. Broxholme, S.F., R.G., E.H., C.H., L.H., P.H., A.K., S. Lise, G.L., D.M., L.M., A. Rimmer, N.S., B.W., C.Y. and N. Popitsch performed study-wide bioinformatic analysis of whole-genome sequence data, led by J.-B.C. and R.R.C. J.T. performed the whole-exome sequence analysis presented in Supplementary Figure 10. E.E.D., A.V.G., M.H., J.L., H.C.M., S.J.M., K.A.M., A.P., L.Q. and P.A.v.S. performed project-specific bioinformatic analysis of whole-genome sequence data. A.R.A., O.C., A.L.F., A. Goriely, I.H.G., A.V.G., R.H., J.L., K.A.M. and A.P. performed project-specific genetic and functional validation studies. G.M. wrote the manuscript with help from H.C.M., J.C.T. and A.O.M.W. and further contributions from S. Lise, D.M., A.P., R.V.T. and S.E.P. J.C. collated information for the paper. P.D. chaired the Steering Committee and the Operations Committee. J.I.B., D. Bentley, G.M., P.J.R., J.C.T. and A.O.M.W. were members of the Steering Committee. J. Broxholme, D. Buck, J.-B.C., R.C., J.C.K., G.L., G.M., J.C.T., I.T., A.O.M.W. and L.W. were members of the Operations Committee.
Corresponding author
Ethics declarations
Competing interests
All authors at Illumina are employees of Illumina, Inc., a public company that develops and markets systems for genetic analysis. G.M., G.L. and P.D. are founders and shareholders of Genomics, Ltd., a company that develops genome analytics.
Integrated supplementary information
Supplementary Figure 1 Distribution of sequencing coverage in the WGS500 project.
Left, plots of the cumulative distribution, for each WGS500 sample, of coverage across the genome (top left) or exome (bottom left). Top right, a comparison of coverage between WGS500 samples (blue) and exomes (black) sequenced at the Oxford Biomedical Research Centre. Thicker lines are the medians across samples; dotted vertical lines are the global medians. Bottom right, the distribution, for each WGS500 sample, of the ratio of the number of reads with the alternate allele (ALT) to the total number of reads (TOTAL), for novel variants. We expect the mean to be 0.5. Individuals with mean <0.4 are shown with colored lines. These are likely to have sample contamination, which leads to a larger number of heterozygous calls for which there are few ALT reads. The sample HCM_2361 was removed from further analysis.
Supplementary Figure 2 Influence of coverage on concordance between sequence data and SNP arrays for multiple samples.
Left, genotype concordance as a function of sequencing depth; note that concordance drops progressively when coverage drops below 15×. 95% confidence intervals, calculated by the Wald method, are indicated. Right, fraction of sites with a given level of coverage. Note that samples with higher coverage (e.g., LVNC_1.1.70, LVNC_1.2.83) have fewer SNPs in the lower-coverage bins, and the genotype concordance estimate therefore has larger confidence intervals.
Supplementary Figure 3 Effect of filtering variants by frequency in public databases and/or other WGS500 samples.
Density plots of the distribution of the number of novel heterozygous (top) or rare homozygous (bottom) coding variants (ANNOVAR annotation) across all individuals, where frequency is defined in the control data sets indicated. The individuals in the top 5th percentile are shown; all of these samples are known to have African or South Asian ancestry except for MR_6 and MR_8, for which we suspect have some sample contamination (Supplementary Fig. 1). ESP, NHLBI Exome Sequencing Project.
Supplementary Figure 4 The burden of variants of unknown significance in candidate genes for craniosynostosis.
Histograms of the number of potentially pathogenic, conserved coding variants in different candidate gene sets for craniosynostosis (CRS). The candidate genes were chosen by a combination of literature and high-throughput database searches, augmented by expert curation (Online Methods). Sample names in green text indicate that the variant is not likely to be pathogenic, as it does not fit a plausible inheritance model or is less functionally compelling than another candidate (Supplementary Table 6). SC, Saethre-Chotzen syndrome.
Supplementary Figure 5 The burden of putative regulatory variants.
Distributions of the number of novel heterozygous (top) and rare homozygous (bottom) variants that alter conserved positions in regulatory regions within 5 kb (red) or 50 kb (black) of a gene. The fact that the number of variants does not substantially change if one considers only regulatory regions within 5 kb of genes (black line) reflects the fact that these regions tend to be close to genes. Note that most of the outliers are also outliers in Supplementary Figure 3, and these samples tend to be of African or Asian ancestry.
Supplementary Figure 6 The burden of putative regulatory variants around candidate genes for early-onset epilepsy or craniosynostosis.
As shown for Supplementary Figure 4 but for variants at conserved positions in regulatory regions within 50 kb of candidate genes for early-onset epilepsy (top) or craniosynostosis (bottom).
Supplementary Figure 7 Segregation of putative causal variants in UMOD and CASR.
Top, the NM_001008389:c.410G>A UMOD variant was identified by WGS in individual III.2 in this family with familial juvenile hyperuricaemic nephropathy (FJHN). The G>A transition generates an AccI restriction endonuclease recognition site, and digestion of a 349-bp PCR product with AccI was used to confirm cosegregation of the variant with affected individuals in the family. Digestion of the mutant (mut) allele generated 93-bp and 256-bp fragments, with the wild-type (WT) allele remaining uncut. Bottom, the NM_000388:c.2299G>C CASR variant was identified by WGS in individuals I.2 and II.1 in a family with familial hypoparathyroidism (FH). The G>C transversion causes loss of a BssSI restriction endonuclease recognition site, and digestion of a 367-bp PCR product with BssSI was used to confirm cosegregation of the variant with affected individuals in the family. Digestion of the wild-type (WT) allele generated 180-bp and 187-bp fragments, with the mutant (mut) allele remaining uncut.
Supplementary Figure 8 Parental origin and sequence conservation of the HUWE1 mutation.
Left, alignment of sequencing reads from the proband (CRS_4659), mother (CRS_4654) and father (CRS_4655) over 2 C/A polymorphisms (arrows; C shown in red and A shown in black). Allele-specific primers (AARev and CCRev) were designed with a common primer Intron6-For to amplify the HUWE1 mutation and polymorphisms in a single PCR product (top right). Red arrows indicate polymorphic sites, and nucleotides included in the primer sequences are underlined. The results of PCR are shown underneath. The products were digested with HpaII, which showed the presence of the mutation (white arrow) only in the CC-Rev amplification product from the proband (second panel from the bottom, right), indicating paternal origin. Bottom right, an alignment of the DUF908 domain in the protein encoded by HUWE1, with the mutated residue indicated (red arrow).
Supplementary Figure 9 Identification of an inherited interstitial insertion involving chromosomes 2p25.3 and Xq27.1 associated with X-linked recessive hypoparathyroidism.
The sequences of the proximal (top left) and distal (top right) insertion junctions are shown. Reference sequences on Xq27.1 and 2p25.3 are indicated in red and blue, respectively. A 3-bp microinsertion at the distal insertion boundary is indicated in yellow. Bottom, primers specific for chromosomes 2 (2SPF) and X (XSPF and XSPR) were designed for the DNA sequence at the distal boundary and used to further characterize the insertion. The sizes of the PCR products obtained with each primer pair are indicated. Chromosome X is shown in black, and the inserted sequence from chromosome 2q25.3 is shown in gray.
Supplementary Figure 10 Coverage in this study compared to a large-scale exome sequencing project.
Coverage comparison of this study and a large-scale exome sequencing (WES) project for the variants given in Table 1 (top) and the causative variants identified in the WES project (bottom). For the WES project, for nondisclosure reasons, only the gene name is given. The WES coverage data (blue) were compiled from 141 whole-exome data sets that were sequenced using the Roche NimbleGen SeqCap EZ v.2.0 kit. Labels for variants located in regions targeted by this kit are in blue, those within 20 bp of the targeted regions are in green and those outside the targeted regions are in red. The WGS500 data (red) were compiled from all the whole-genome data sets used in this study. The horizontal green lines denote two exemplary coverage thresholds used in variant detection. To improve readability, the plots were truncated above a coverage value of 100 (top) or 200 (bottom) and the box-plot whiskers were extended to the data extremes.
Supplementary Figure 11 Distribution of the lengths of the largest regions of homozygosity across all samples.
Thirty-seven samples had at least one region of homozygosity >4 Mb in length (black bars), suggesting consanguinity. Note that the largest bin includes one sample with confirmed uniparental isodisomy. See the Online Methods for an explanation of how regions of homozygosity were identified.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–11, Supplementary Tables 1–10 and Supplementary Note. (PDF 1908 kb)
Rights and permissions
About this article

Cite this article
Taylor, J., Martin, H., Lise, S. et al. Factors influencing success of clinical genome sequencing across a broad spectrum of disorders. Nat Genet 47, 717–726 (2015). https://doi.org/10.1038/ng.3304
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ng.3304
This article is cited by
-
PDIVAS: Pathogenicity predictor for Deep-Intronic Variants causing Aberrant Splicing
BMC Genomics (2023)
-
Whole genome sequencing diagnostic yield for paediatric patients with suspected genetic disorders: systematic review, meta-analysis, and GRADE assessment
Archives of Public Health (2023)
-
Monogenic inflammatory bowel disease-genetic variants, functional mechanisms and personalised medicine in clinical practice
Human Genetics (2023)
-
Structural and non-coding variants increase the diagnostic yield of clinical whole genome sequencing for rare diseases
Genome Medicine (2023)
-
Whole exome analysis of patients in Japan with hearing loss reveals high heterogeneity among responsible and novel candidate genes
Orphanet Journal of Rare Diseases (2022)
