
Abstract
Coral growth rates are highly dependent on environmental variables such as sea surface temperature1,2 and solar irradiance3,4. Multi-decadal variability in coral growth rates has been documented throughout the Caribbean over the past 150–200 years5,6, and linked to variations in Atlantic sea surface temperatures5. Multi-decadal variability in sea surface temperatures in the North Atlantic, in turn, has been linked to volcanic and anthropogenic aerosol forcing7. Here, we examine the drivers of changes in coral growth rates in the western Caribbean between 1880 and 2000, using previously published coral growth chronologies from two sites in the region, and a numerical model. Changes in coral growth rates over this period coincided with variations in sea surface temperature and incoming short-wave radiation. Our model simulations show that variations in the concentration of anthropogenic aerosols caused variations in sea surface temperature and incoming radiation in the second half of the twentieth century. Before this, variations in volcanic aerosols may have played a more important role. With the exception of extreme mass bleaching events, we suggest that neither climate change from greenhouse-gas emissions nor ocean acidification is necessarily the driver of multi-decadal variations in growth rates at some Caribbean locations. Rather, the cause may be regional climate change due to volcanic and anthropogenic aerosol emissions.
Similar content being viewed by others

Main
Coral reefs have been under increasing pressure as a result of multiple anthropogenic stressors8, especially in the Caribbean9. Nevertheless, attributing past changes in coral reef ecosystems to specific anthropogenic activities such as greenhouse-gas (GHG) and aerosol emissions remains difficult10, contributing to large uncertainty in modelling future climatic impacts on coral reefs.
Coral growth rates (extension rates) are a useful indicator of ecosystem health11 and are highly responsive to climatically influenced environmental variables such as solar irradiance3,4 and sea surface temperatures1,2 (SST). Long-term (decadal and longer) variability in Atlantic SSTs, termed the Atlantic Multi-decadal Oscillation12 (AMO), has been shown to influence the corals of this region. Caribbean coral skeletal δ18O signatures correlate with the AMO (ref. 13) and in the northern Caribbean it has been suggested that corals contain multi-decadal oscillations in growth rates that are negatively correlated with the AMO (refs 5, 14).
Until recently, the prevailing consensus has been that the AMO is a mode of internal variability within the climate system15. However, there is increasing evidence pointing towards volcanoes and anthropogenic aerosols as drivers of AMO variability during the observed period. Large volcanic events are linked to cool phases in SSTs (ref. 16) and recent work revisiting Atlantic variability in present global climate models also identifies an important role of past anthropogenic aerosol emissions in determining much of the observed multi-decadal change7,16,17. Aerosols influence SSTs by altering solar (short-wave) radiation fluxes. We look to extend the perspective of previous authors7 and examine the dual ecosystem impacts of aerosol-driven SST and short-wave radiation variability. Corals provide an ideal candidate for such an analysis because their calcification is driven by both factors and the outcome is preserved in long-lived coral skeletons.
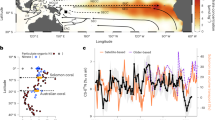
We analysed previously published continuous coral growth chronologies from two sites in the western Caribbean, Belize18 (17.50° N, 87.76° W) and Panama6 (9.25° N, 79.50–80.00° W; Fig. 1). The Belize data set used was the Turneffe atollMontastraea faveolata master chronology from 1905 to 1998 (ref. 18). The Panama data set used was the Siderastrea siderea master chronology from 1880 to 1989 (ref. 6; further details in Supplementary Information). A 13-year filter was applied to the growth anomalies to highlight multi-decadal variability, following the methodology of previous studies for example19 and a Bayesian hierarchical model was used to infer the significance of potential physical predictors.

Mean SST map of the Greater Caribbean region showing the Belize (i) and Panama (ii) sites. Contours give the standard deviation of the mean. SSTs are mean values from the Operational Sea Surface Temperature and Sea Ice Analysis (OSTIA) high-resolution satellite data set (1985–2007)30.
Coral cores from the sites we selected show multi-decadal signals in growth rates that have periodicity synchronous with the AMO (Fig. 2). We brought together the available observational data sets with output from the Earth system model HadGEM2-ES (ref. 20) run with time-varying and pre-industrially fixed anthropogenic aerosol emissions. Three potential physical predictors of coral growth rate (SST (refs 1, 2), short-wave radiation3,4 and aragonite saturation state3,4) were systematically tested for statistically robust relationships with coral growth rates. We found that changes in coral growth rates are consistent with the hypothesis that such variability is driven by SST and short-wave radiation change (Table 1). Aerosol particles both scatter incoming radiation, and act as nucleation sites for cloud droplets. With increasing aerosol concentrations, here primarily in the form of SO4, there is increased direct scattering of short-wave radiation from aerosol particles, and more, smaller, cloud-droplets, resulting in more reflective and longer-lasting clouds. These factors reduce the net downwelling short-wave radiation reaching the sea surface. This reduces the photosynthetically active radiation that corals and their algal symbionts receive as well as that received in waters subsequently transported laterally into the region by the local circulation and mixing. This in turn affects SST, affecting coral metabolic rate and calcification within a certain range4,5,21. Observed SST anomalies are positively correlated with multi-decadal coral growth anomalies in Belize and Panama (Fig. 2b,c). The lack of exact phase synchrony between coral growth anomalies and observed SST anomalies is most likely due to the influence of other localized natural and anthropogenic drivers such as human disturbance, changes in runoff and potential nutrient enrichment. This is especially the case at the Panama site in the mid-twentieth century where coastal industrial activity is known to have impacted coral growth6. It is also one of the Caribbean regions with the highest runoff22. Both changes in salinity and sediment load/turbidity have the potential to influence coral growth23.

a, The 25-year filtered AMO. b,c, 25-year filtered coral growth anomalies (blue) and observed SST anomalies (red) for the Belize (b) and Panama (c) sites. Coral growth and SST are linearly detrended as per the definition of the AMO.
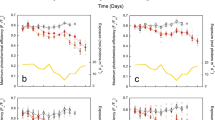
Aerosols were recently shown to be of critical importance in driving past multi-decadal oscillations in North Atlantic SSTs (ref. 7). HadGEM2-ES accounts for both the long-term increase and multi-decadal variability in Caribbean average SST (Fig. 3a). When run with anthropogenic aerosol emissions fixed at pre-industrial values, HadGEM2-ES fails to recreate multi-decadal SST variability in the mid-to-late twentieth century, especially post 1960 (Fig. 3a). The marked 1960–1970 SST decrease is thought to be the result of rising anthropogenic aerosol emissions associated with post-World War II industrial expansion in North America and to a lesser extent Central and South America7. Around 1970 when the introduction of clean-air policies markedly reduced aerosol emissions, SSTs stabilize and subsequently begin to rise again in both the model output with time-varying anthropogenic aerosol emissions and the observational record (Fig. 3a).

Modelled SST and short-wave radiation (SW) with time-varying (blue) and fixed (red) anthropogenic aerosols. a,b, SST is shown as Caribbean average (a) and short-wave radiation is shown for the Belize and Panama sites (b). Model ensemble uncertainty is shown bounded by dashed lines and variables are 13-year filtered. Observed Caribbean average SST is shown in black and modelled SSTs have been bias corrected. See Methods for further details on modelling methodology.
One process HadGEM2-ES does not capture is the positive feedback from dust on Atlantic SST change (cool SSTs lead to drier African conditions and more dust, which have been linked to Atlantic short-wave radiation and SST reductions24). HadGEM2-ES captures annual variability, but not the multi-decadal7, observed in the eastern Caribbean25. This is one explanation for why modelled Caribbean average SST does not precisely capture the observed SST variability (Fig. 3a).
When run with time-varying and fixed anthropogenic aerosol emissions, HadGEM2-ES short-wave radiation shows significant differences at the Belize and Panama sites (Fig. 3b). The influence of anthropogenic aerosols on short-wave radiation at the Belize site is substantial. That is, there is limited influence before 1960, after which aerosols seem to have a strengthening role in reducing short-wave radiation. This is not the case at the Panama site, where anthropogenic aerosols seem to have less influence on short-wave radiation throughout the historical period (Fig. 3b). Such differences are probably due to the limited atmospheric dispersal of aerosols and the greater proximity of the Belize site to the US Eastern Seaboard (the predominant source of twentieth-century anthropogenic aerosol emissions in the region) as well as differing climatic regimes (see Supplementary Information).
To account for auto-correlation in the filtered coral growth time series we apply the following statistical model, which allows for temporal dependence but also the incorporation of predictors:

where SW is short wave radiation. This is a simple state-space model as successfully applied to other environmental issues26 and is effectively a conditional normal linear model with a time-dependent intercept. The intercept β0(t) is assumed random, where the present value is centred on the previous one. The random intercept accounts for auto-correlation in the response y(t) while allowing for predictor effects to be assessed through the significance in βi. To prevent β0(t) from over-smoothing the data the variance τ2 is restricted so that τ2/σ2 is in the range [0, 1].
The three potential physical predictors of coral growth rate were HadGEM2-ES Caribbean average SST, site-specific short-wave radiation and aragonite saturation state (Ωarag). To select the predictors in the statistical models of the two sites, the deviance information criterion (DIC) was used. The procedure for model selection involved starting with a null model y(t) = β0(t)+ɛ(t) and adding predictors systematically to minimize the DIC. The optimal model produced by this methodology does not necessarily contain all predictors.
The final model for Panama included only SST and short-wave radiation whereas for Belize, all three predictors were included. As the models were Bayesian, inference on each parameter is based on the posterior distribution of each of them. Significance is assessed by looking at the 95% confidence interval of each parameter. If the value of zero is in the tails of the posterior distribution, this indicates significance as there is more certainty that the parameter is either positive or negative. As the confidence intervals of both β1 and β2 of the Belize model span positive values and do not contain zero, SST and short-wave radiation have a positive strongly significant effect on coral growth. Parameter β3 relating to Ωarag has a negative effect that is mildly significant (Table 1). SST in the Panama model is not very influential as the confidence interval for β1 is mostly positive but has considerable mass on the negative side. There is more confidence that short-wave radiation has a negative effect as most of the posterior distribution of β2 is negative (Table 1). It is important to note that although the SST and short-wave radiation regression parameters for Panama are significant (according to the DIC and 95% confidence intervals) they are not as conclusive as for Belize.
The mildly significant negative Ωarag estimate β3 is contrary to knowledge of its influence on coral growth4,5. This is probably a combined result of the long-term upward trend in coral growth rates at the Belize site (Supplementary Fig. S1) and the near-linear decline of aragonite saturation state over the historical period (Supplementary Fig. S2). We therefore suspect that Ωarag is acting as a surrogate for time and its significance is spurious. This limited role of aragonite saturation state in explaining historical coral growth rate variability is in line with recent findings in Florida14 and Western Australia27. It should also be noted that the relationship between offshore Ωarag as produced by HadGEM2-ES and inshore Ωarag as experienced by many coral reefs is dependent on coastal processes and the residence time of water28. At the more inshore location of the Panama site, long-term changes in Ωarag are potentially less likely to reflect HadGEM2-ES model values, although observational data sets to test such a hypothesis do not exist.
When the models are forced by predictors derived from fixed anthropogenic aerosol emission runs, they still capture the late nineteenth century and early twentieth century variability in growth rates that is thought to be largely driven by volcanic activity7 (Fig. 4). However, post-1950 in Belize and post-1970 in Panama these models increasingly fail to capture observational coral growth rates and diverge from the all-forcings models because of the greater role of anthropogenic aerosols in this period (Fig. 4). Across both sites, but especially at the Belize site, anthropogenic aerosols seem to have suppressed coral growth rates as a result of aerosol-driven effects on short-wave radiation and SST. The extent of divergence between models using fixed and time-varying anthropogenic aerosol emissions is far greater for Belize than Panama (Fig. 4). This is in part due to the physical predictors for Belize being stronger and anthropogenic aerosols having less influence on Panama short-wave radiation (Fig. 3b).

a,b, Observational coral growth (black), and modelled coral growth with time-varying anthropogenic aerosols (blue) and with fixed anthropogenic aerosols (red) for the Belize (a) and Panama (b) sites. The 95% confidence intervals of the regression model with time-varying and fixed anthropogenic aerosols are shown bounded by dashed lines. This uncertainty includes HadGEM2-ES ensemble uncertainty. Observations and model variables are 13-year filtered.
The previous correlations that have been found between coral growth and the AMO are now resolved at two sites in the western Caribbean by considering the proximate drivers of light and SST directly. The conclusion that past changes in coral growth rates are consistent with changes in atmospheric aerosol concentration has strong implications for the ecosystem impacts of increasingly stringent clean-air legislation, industrialization in the developing world and potential aerosol-based geoengineering. As such, coral ecosystems are likely to be sensitive to not only the future global atmospheric CO2 concentration but also the regional aerosol emissions associated with industrialization and decarbonization. Accurate projections of future coral growth will thus need to take into account, not just future GHG concentrations, but also emission pathways of aerosol precursors.
Methods
Coral analysis.
The Belize data set used was the Turneffe atoll Montastraea faveolata master chronology from 1905 to 1998 comprised of 9 cores18 (17.50° N, 87.76° W). Core slabs (0.86 cm) were X-rayed using a Siemens Polyphos 50 with a source-to-object distance of 40 inches and a setting of 63 kV at 5 mA s−1. Coral X-rays were analysed for annual linear extension, using CoralXDS and the second-derivative zero-band delimiting function to objectively identify the beginning and end of each band. For each coral core, three transects at various locations on the core were analysed and averaged to account for slight variations in within-band extension. The Panama data set used was the entire Bahia las Minas Siderastrea siderea master chronology from 1880 to 1989 comprised of 77 cores6 (9.25° N, 79.50°–80.00° W). Standard methods were used to process the cores and construct the master chronology. Annual growth extension rates were measured from the previous year’s outermost high-density band from each colony using a Sigma-Scan 3.90 digitizer at a 0.001-mm resolution. Annual growth patterns were previously calibrated for the locality (further details in Supplementary Information).
Environmental records.
Observational SST was obtained from the 1°×1° gridded Hadley Centre Sea Ice and Sea Surface Temperature (HadISST 1.1) data set (1870–2006). The HadISST 1.1 data set consists of blended in situ observations and satellite Advanced Very High Resolution Radiometer (AVHRR) measurements29. Modelled SST, solar irradiance and aragonite saturation state were obtained from the gridded Hadley Global Environment Model 2-Earth System (HadGEM2-ES) outputs (1870–2005). Within HadGEM2-ES, aragonite saturation state (Supplementary Fig. S2) is calculated from dissolved carbon and alkalinity concentrations, temperature, salinity and assuming the calcium concentration to be in a fixed ratio with salinity. The physical and chemical fields used within the carbonate chemistry calculation are all simulated interactively within the HadGEM2-ES model; the CO2 partial pressure calculated contemporaneously with the saturation state has been shown to validate well at the Bermuda Atlantic Time-Series, and spatially against observations. Model resolution increases from 1°×1° in high latitudes to 1/3°×1/3° at the Equator. Means were taken across ensemble members (4 for the historical scenario and 3 for the scenario with fixed anthropogenic aerosol emissions). The AMO index was constructed from the HadISST SST data set.
Earth system model simulations.
HadGEM2-ES models the supply of oxidants, an important component for aerosol formation, and mineral dust aerosols interactively and includes biomass and carboniferous aerosol properties. Source terms for natural aerosols (or precursors) and mineral dust are also modelled interactively. The ensemble of 4 historical simulations is forced with driving data (GHGs, aerosols, volcanoes and solar changes) based on historical data sets compiled for Coupled Model Intercomparison Project Phase 5 (CMIP5) simulations. Volcanic forcing is prescribed in latitudinal bands. Over the North Atlantic the magnitude of optical depth changes is prescribed individually for 0°–30° N, and 30°–90° N, capturing the differences in tropical and extra-tropical volcanoes. Individual members were initiated from a control simulation using start points located 50 years apart.
The ensemble of 3 HadGEM2-ES simulations with fixed anthropogenic aerosol emissions used identical driving data to the standard historical ensemble, prescribing changes in emissions and concentrations based on the CMIP5 historical data sets. The exception is the anthropogenic aerosol emissions and surface chemistry (and consequent contribution to aerosol oxidation), which were kept constant at their year 1860 values. This ensemble provides a comparison of historical SST and short-wave radiation where the historical changes in anthropogenic aerosol emissions did not take place.
Modelling.
Global Earth system models contain neither the relevant processes, nor physical resolution to accurately simulate coastal ocean conditions; consequently, modelled Caribbean average, rather than local SSTs, were used in statistical models. Owing to the lack of observational data, it is not possible to assess the extent to which localized solar irradiance is historically representative of that observed at a regional scale. However, the limited atmospheric dispersal and lifetime of aerosols would suggest that coherence between regional and local irradiance in the Caribbean is unlikely. We therefore chose to use localized irradiance outputs in our models.
The statistical modelling was implemented in a Bayesian framework using the freely available statistical software R (http://www.R-project.org/) and WinBUGS (http://www.mrc-bsu.cam.ac.uk/bugs). As the statistical model intercept β0(t) is assumed random but unobserved, the resulting models are able to capture complex forms of time dependence in y(t).
Filtering.
A 13-year filter was applied to SST, solar irradiance and coral growth data sets to remove short-term interannual variability before constructing linear models. The filter period was chosen on the basis of available coral core data. Previous studies analysing AMO-driven ecological changes have adopted a similar filtering approach to dampen the effect of short-timescale interannual variability19.
References
Lough, J. & Barnes, D. Environmental controls on growth of the massive coral Porites. J. Exp. Mar. Biol. Ecol. 245, 225–243 (2000).
Lough, J. & Barnes, D. Several centuries of variation in skeletal extension, density and calcification in massive Porites colonies from the Great Barrier Reef: A proxy for seawater temperature and a background of variability against which to identify unnatural change. J. Exp. Mar. Biol. Ecol. 211, 29–67 (1997).
Langdon, C. & Atkinson, M. Effect of elevated p CO 2 on photosynthesis and calcification of corals and interactions with seasonal change in temperature/ irradiance and nutrient enrichment. J. Geophys. Res. 110, C09S07 (2005).
Marubini, F., Barnett, H., Langdon, C. & Atkinson, M. Dependence of calcification on light and carbonate ion concentration for the hermatypic coral Porites compressa. Mar. Ecol. Prog. Ser. 220, 153–162 (2001).
Saenger, C., Cohen, A., Oppo, D., Halley, R. & Carilli, J. Surface temperature trends and variability in the low-latitude North Atlantic since 1552. Nature Geosci. 2, 492–495 (2009).
Guzman, H., Cipriani, R. & Jackson, J. Historical decline in coral reef growth after the Panama Canal. Ambio 37, 342–346 (2008).
Booth, B., Dunstone, N., Halloran, P., Andrews, T. & Bellouin, N. Aerosols implicated as a prime driver of 20th century North Atlantic climate variability. Nature 484, 228–232 (2012).
Hughes, T. et al. Climate change, human impacts, and the resilience of coral reefs. Science 301, 929–933 (2003).
Pandolfi, J. et al. Global trajectories of the long-term decline of coral reef ecosystems. Science 301, 955–958 (2003).
Hoegh-Guldberg, O. Low coral cover in a high-CO2 world. J. Geophys. Res. 110, C09S06 (2005).
Smith, S. & Buddemeier, R. Global change and coral reef ecosystems. Annu. Rev. Ecol. Syst. 23, 89–118 (1992).
Kerr, R. A North Atlantic climate pacemaker for the centuries. Science 288, 1984–1985 (2000).
Hetzinger, S. et al. Caribbean coral tracks Atlantic Multidecadal Oscillation and past hurricane activity. Geology 36, 11–14 (2008).
Helmle, K., Dodge, R., Swart, P., Gledhill, D. & Eakin, C. Growth rates of Florida corals from 1937 to 1996 and their response to climate change. Nature Commun. 2, 215 (2011).
Knight, J. The Atlantic multidecadal oscillation inferred from the forced climate response in coupled general circulation models. J. Clim. 22, 1610–1625 (2009).
Ottera, O., Bentsen, M., Drange, H. & Suo, L. External forcing as a metronome for Atlantic multidecadal variability. Nature Geosci. 3, 688–694 (2010).
Chang, C., Chiang, J., Wehner, M., Friedman, A. & Ruedy, R. Sulfate aerosol control of tropical Atlantic climate over the twentieth century. J. Clim. 24, 2540–2555 (2011).
Carilli, J., Norris, R., Black, B., Walsh, S. & McField, M. Century scale records of coral growth rates indicate that local stressors reduce coral thermal tolerance threshold. Glob. Change Biol. 16, 1247–1257 (2010).
Martinez, E., Antoine, D., D’Ortenzio, F. & Gentili, B. Climate-driven basin-scale decadal oscillations of oceanic phytoplankton. Science 326, 1253–1256 (2009).
Collins, W. et al. Development and evaluation of an earth-system model—HadGEM2. Geosci. Model Dev. Discuss. 4, 997–1062 (2011).
Gill, J., Watkinson, A., McWilliams, J. & Côté, I. Opposing forces of aerosol cooling and El Niño drive coral bleaching on Caribbean reefs. Proc. Natl Acad. Sci. USA 103, 18870–18873 (2006).
Chollett, I., Mumby, P. J., Müller-Karger, F. E. & Hu, C. Physical environments of the Caribbean Sea. Limnol. Oceanogr. 57, 1233–1244 (2012).
Fabricius, K. E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 50, 125–146 (2005).
Evan, A. T., Foltz, G. R., Zhang, D. & Vimont, D. J. Influence of African dust on ocean-atmosphere variability in the tropical Atlantic. Nature Geosci. 4, 762–765 (2011).
Prospero, J. M. & Lamb, P. J. African droughts and dust transport to the Caribbean: Climate change implications. Science 302, 1024–1027 (2003).
Sahu, S. K., Gelfand, A. E. & Holland, D. M. High-resolution space-time ozone modelling for assessing trends. J. Am. Statist. Assoc. 102, 1221–1234 (2007).
Cooper, T., OLeary, R. & Lough, J. Growth of Western Australian corals in the Anthropocene. Science 335, 593–596 (2012).
Anthony, K., Kleypas, J. A. & Gattuso, J. P. Coral reefs modify their seawater carbon chemistry-implications for impacts of ocean acidification. Glob. Change Biol. 17, 3655–3666 (2011).
Rayner, N. et al. Global analyses of sea surface temperature, sea ice, and night marine air temperature since the late nineteenth century. J. Geophys. Res. 108, 4407 (2003).
Donlon, C. J. et al. The Operational Sea Surface Temperature and Sea Ice analysis (OSTIA). Remote Sens. Environ. 116, 140–158 (2012).
Acknowledgements
We thank R. Williams, C. Luke, A. Hunter, D. Long, C. Saenger, J. P. Carricart-Ganivet and R. Iglesias-Prieto for helpful insight. We also thank J. Hughes for undertaking the all-forcings model simulations. The study was financially supported by a NERC grant to P.J.M. and P.M.C., the University of Exeter and the EU FORCE project. P.R.H. was supported by the Joint DECC/Defra Met Office Hadley Centre Climate Programme (GA01101).
Author information
Authors and Affiliations
Contributions
L.K., P.M.C. and P.R.H. conceived and designed the experiments. L.K. performed the experiments and analysed the data. L.K., P.M.C., P.R.H., T.E., P.J.M., J.C., H.M.G. and B.B.B.B. interpreted the data, discussed their implications and contributed to the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Information
Supplementary Information (PDF 549 kb)
Rights and permissions
About this article
Cite this article
Kwiatkowski, L., Cox, P., Economou, T. et al. Caribbean coral growth influenced by anthropogenic aerosol emissions. Nature Geosci 6, 362–366 (2013). https://doi.org/10.1038/ngeo1780
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ngeo1780
This article is cited by
-
Coral extension rate analysis using computed axial tomography
Coral Reefs (2022)
-
The formation of biogenic reef stone: from coral skeleton to reef rubble
Journal of Oceanography (2022)
-
Coral bleaching under unconventional scenarios of climate warming and ocean acidification
Nature Climate Change (2015)
-
Historic impact of watershed change and sedimentation to reefs along west-central Guam
Coral Reefs (2014)
-
What spatial scales are believable for climate model projections of sea surface temperature?
Climate Dynamics (2014)
