
Abstract
Testing declarative memory in laboratory rodents can provide insights into the fundamental mechanisms underlying this type of learning and memory processing, and these insights are likely to be applicable to humans. Here we provide a detailed description of the social discrimination procedure used to investigate recognition memory in rats and mice, as established during the last 20 years in our laboratory. The test is based on the use of olfactory signals for social communication in rodents; this involves a direct encounter between conspecifics, during which the investigatory behavior of the experimental subject serves as an index for learning and memory performance. The procedure is inexpensive, fast and very reliable, but it requires well-trained human observers. We include recent modifications to the procedure that allow memory extinction to be investigated by retroactive and proactive interference, and that enable the dissociated analysis of the central nervous processing of the volatile fraction of an individual's olfactory signature. Depending on the memory retention interval under study (short-term memory, intermediate-term memory, long-term memory or long-lasting memory), the protocol takes ∼10 min or up to several days to complete.
Similar content being viewed by others
Introduction
Soon after the beginning of the systematic investigation of human memory in the nineteenth century, scientists classified different types of memory. One of the pioneers in this field, the German psychologist Hermann Ebbinghaus, classified not only differences in the duration of the ability to recall a learned item but also whether conscious or unconscious access to the memory is required1. Today, memory whose recall requires unconscious access to the learned items is called nondeclarative or implicit memory2. Memory processes characterized by a conscious recall are called declarative or explicit memory.
Characteristics of recognition memory
A typical example of a declarative memory process is recognition memory, which refers to the ability both to separate familiar from unfamiliar items and to identify these items3. Testing recognition memory in the laboratory requires two sessions (Fig. 1a): during the first ('learning') session, the item to be recognized is shown to the experimental subject to allow the acquisition ('sampling') of the relevant information. The item is then removed, thus initiating an interval (the 'retention interval') during which the information obtained during 'sampling' is stored by the experimental subject. Retention of the memory is then tested in a 'choice' session during which the experimental subject is simultaneously exposed to the item presented during 'sampling' and to a similar, novel item that has not previously been encountered. By increasing the interval between 'sampling' and 'choice', retention performance can be tested, ranking from short-term (minutes to several hours), intermediate-term (8–12 h) and long-term (18 h to a few days) up to long-lasting memory (>7 d)3.

(a) Experimental sessions for testing recognition memory. During a 'sampling' session, the 'to be recognized' item is presented; after a fixed retention interval, there is a 'choice' session, in which the item is re-exposed to the experimental subject together with a previously unencountered item. (b) Illustration of the social discrimination procedure in laboratory rats and mice. The experimental subject (Es; an adult male or female animal) is exposed in its home cage to conspecific stimulus animals, in this case, juveniles (J) during both 'sampling' and 'choice'. Each session lasts 4 min, and they are separated by a fixed interval that (in case of successful recognition during 'choice') corresponds to the retention interval.
Test paradigms to study recognition memory
The investigation of recognition memory is a prospering field in psychology that has produced a vast literature of human studies. However, unraveling the neurobiological mechanisms underlying recognition memory, including inter- and intracellular signaling networks within distinct brain areas, requires animal experimentation. Over the last 20 years, procedures have been established that allow recognition memory to be studied in the two most common laboratory rodents: rats and mice. In many of these procedures, an additional component—either rewarding or punishing—is introduced to allow an easier readout of the behavioral performance that is used as an index for memory. However, the presentation of such additional, 'to be associated', stimuli may limit the extrapolation of animal data concerning the neuronal mechanisms that underlie recognition memory.
There are two obvious problems. First, processing of an additional stimulus is stimulated (e.g., if food or a sweet fluid is provided as a reward, there is gustatory stimulus processing). As a consequence, a substantial proportion of neuronal activity is linked to (i) the acquisition and storage of recognition memory and (ii) the processing of the additional stimulus, which may itself reach 'memory' quality (e.g., ref. 4); it is also required for stimulus-stimulus association. Second, presenting a rewarding or punishing stimulus adds an additional emotional component (reinforcer) to the item to be remembered. Such reinforcers can cause a higher resistance of memory against forgetting5. Indeed, at least for fear conditioning, a decline in the behavioral parameters used to measure 'extinction' in paradigms on the basis of stimulus-stimulus associations may not simply result from a weakening of the association strength but may be due to habituation toward the conditioned stimulus6. In other words, once a stimulus-stimulus association has been learned, the experimental subject will not respond again to that conditioned stimulus in the same way as a naive subject would. As a consequence, the same animal cannot be exposed to the same combination of stimuli under different treatment conditions. Instead, the performance of separate groups of animals (e.g., control versus treatment) must be compared statistically.
As a way of avoiding these problems, there has been increasing interest in nonconditioned procedures that allow the investigation of recognition memory in laboratory rodents. These tests rely on the spontaneous investigatory behavior of the animals, and on the fact that nonfamiliar items will be investigated more than familiar items. The complex social behavior of rats and mice requires them to be able to recognize familiar conspecifics; taking advantage of this, social recognition testing seems to provide a unique approach to recognition memory in these species. Animals can be tested repeatedly in social recognition memory procedures without the problem of 'highly persistent memory traces'; thus, the same animals can be tested under different treatment conditions (e.g., treated and untreated or treated with verum versus placebo). This means that relatively few animals are needed to achieve a high statistical power in data analysis.
The social recognition memory task described below exploits these advantages; it is also inexpensive and does not require any special apparatus or expensive software. Moreover, analyzing social behavior provides a sensitive indicator of the general well-being of laboratory rodents7. This is particularly important, as it allows the detection of side effects that may affect the animals' behavior, for instance, in the context of genetic or pharmacological manipulations.
Overview of the nonconditioned social discrimination test
Tests of social recognition evolved from the empirical study of Thor and Holloway8 were introduced by Dantzer and co-workers9 as routine laboratory procedures and were progressively modified and refined to become the 'social discrimination test' as it is known today10.
The test consists of two 4-min exposures of juveniles (stimulus animals) to an experimental subject in the subject's cage. During 'sampling', a stimulus animal is exposed to the experimental subject. The stimulus animal is then removed and kept individually in a fresh cage with food and water ad libitum. After a certain fixed interval, this stimulus animal is re-exposed to the experimental subject together with an additional, novel stimulus animal ('choice' session).
The time spent by the experimental subject in investigating each stimulus animal is measured by a trained observer blind to the animal's treatment. If the experimental subject spends longer investigating the novel stimulus animal than the familiar stimulus animal, this is taken as evidence for an intact recognition memory10. During investigation, the experimental subject acquires the 'olfactory signatures' of the stimulus animals; these consist of both volatile and nonvolatile fractions. Figure 1b shows the olfactory cued social discrimination procedure schematically. Interestingly, short-term social recognition memory seems to be unaffected by both the circadian rhythm in rats11 and chronic stress in CD-1 mice12.
Recently, modifications of this task were proposed, which share the principle that the stimulus animals are separated from the experimental subjects by enclosing them in small cages13,14. This makes 'preference' measurements easier, but it limits access to the nonvolatile fraction of the individual's olfactory signature. It is, therefore, of limited suitability for testing rats in particular; rats need direct access to the conspecific's body surface (primarily the ano-genital region) to show a proper social memory performance15,16,17.
Separating stimulus animals into small cages enables video tracking systems and semiautomated behavioral analysis to be used13,18. However, there are good arguments for direct monitoring of the animals by an experienced observer unaware of the animals' treatment. For example, the olfactory cues that are acquired by the experimental subjects require active sniffing behavior8. This (given the limitations of present technology) can be judged reliably only by a human observer who is close enough to see the breathing movement of the animal directly. It has been repeatedly suggested that presenting the stimulus animals in small enclosures lessens the risk of the experimental subjects scent-marking the stimulus animals14,19. However, there is convincing experimental evidence that, under the experimental conditions described below, (i) this type of scent marking is unlikely to occur, and (ii) even if it does occur, it does not substantially contribute to recognition17,20.
Here we describe an inexpensive, detailed protocol that allows the experimental animals to directly access both the volatile and nonvolatile fractions of the conspecific's olfactory signature16 for testing recognition memory. The advantages and disadvantages of this protocol are summarized in Table 1.
Applications
The test is suitable for investigating the relevance of defined brain areas and distinct neurotransmitter systems involved in short-term to long-lasting social recognition memory. It also makes it possible to investigate the interaction of two similar 'memory traces' during learning, memory consolidation and retrieval20. Tools that have been successfully applied in these studies include permanent lesions of distinct brain areas21, both systemic22,23 and local, intracerebral injections24,25,26,27,28 of substances affecting interneuronal signaling as well as (engineered) mutations29. Again, a particular advantage of the test is that the same animals can be used under treatment and untreated and/or control conditions (e.g., injections with verum and placebo), thereby increasing the statistical power.
Experimental design
When designing an experiment the following aspects should be considered:
Experimental subjects. With respect to short-term versus long-term memory, there are no differences between the sexes in either species. Mice show a memory performance that lasts at least 24 h (refs. 20,22,30) (Supplementary Fig. 1), whereas rats only form a short-term social recognition memory. A more detailed analysis in male rats revealed recognition memory to be extinct after ∼45 min, whereas female rats show a slightly, but significantly longer recognition (∼2 h)9,31,32,33,34. Available data indicate that the recognition abilities of female rats are not affected by the ovarian cycle33 (Supplementary Fig. 1; but see opposite findings for a modified version of the test using sedated stimulus animals and repeated 'sampling'35). We expect that there may be some subtle sex differences in the retention of social recognition memory in mice, but so far this has not been studied. It is notable that in older rats (12, 18 and 24 months) and mice (12–14 months), short-term social memory performance is impaired36. Furthermore, the duration of investigatory behavior during the exposures might be substantially reduced in aged or older animals37 (but see ref. 36).
We (and others) failed to observe differences between standard strains within a given species. In particular, mice of the 129S1SV strain, which show significant differences to other strains in many behavioral parameters38,39,40,41,42,43, do not differ in their social recognition memory performance when tested in the social discrimination task (J.H. and M.E., unpublished observations).
Stimulus animals. Despite one contrary report in the literature44, extensive testing has shown that the sex of juveniles used as stimulus animals has no significant effect on the performance of the experimental subjects in social recognition tests in mice (Fig. 2).

(a,b) Independently of the gender of the juvenile (J) that was exposed during 'sampling' and/or 'choice', adult male mice (for a retention interval (RI) of 24 h; (a)) and rats (for a retention interval of 30 min; (b), except for one protocol: left panel) recognized the juvenile during 'choice' (curly bracket). Statistics was performed by paired t-test for each 'choice' session separately. Juveniles used were of both sexes and aged between 27 and 34 d. For more details see Figure 7.
Retention intervals. Rats show only short-term recognition memory for conspecifics. To measure recognition, most studies have used an exposure interval of 30 min. In contrast, to show memory extinction an exposure interval of 120 min (in male rats) and/or 180 min (in female rats) is used. The results are not affected by the housing conditions of the experimental subjects (group housing versus single housing)16.
Under single-housing conditions, male mice show short-term recognition memory that is usually tested at a retention interval of 60 min; memory extinction is tested at an exposure interval of 120 min45. Group-housed male30 and female mice (Supplementary Fig. 1) show long-term memory for at least 24 h. In our laboratory, a retention interval of 48 h was successfully tested; Kogan et al.30 showed long-lasting social recognition memory for 7 d in group-housed male mice. To study long-term memory, the standard retention interval is 24 h; an exposure interval for the extinction of social memory in group-housed mice is not described.
Effect of cage transfer and anesthesia. Transferring experimental subjects into new cages 10 min both before 'sampling' and 'choice' in adult male C57BL/6JOlaHsd mice had a distracting impact on long-term recognition (Fig. 3a, middle). Moving the experimental subjects 3 h after 'sampling' to a new cage for 4 min and then returning them to their home cage to continue testing also had a distracting impact (Fig. 3a, right). However, moving the experimental subject in its home cage to an adjacent room for 1–4 min and performing the 'sampling' session immediately after returning to the testing room did not affect the recognition performance measured 24 h later (Fig. 3b, left). The same holds true if, during the short sojourn time in the adjacent room, the experimental subjects were removed from the home cage and a short-term lasting (∼2–3 min) isoflurane anesthesia was applied (Fig. 3b, right; see also20,22,23). This makes the social discrimination procedure an interesting tool for testing the impact of pharmacological manipulations on memory performance. Indeed, strains bearing spontaneous34,46 or engineered mutations29 might show impaired memory performance in this task, which can be rescued after appropriate treatment.

(a) Placing the adult male mice as experimental subjects into a novel cage for 4 min. In contrast to their performance when kept without disturbance in their home cages (left), the experimental subjects failed to recognize a juvenile (J) encountered during 'sampling' when transferred immediately before 'sampling' and 'choice' in a new cage (middle) or when, during the retention interval of 24 h, the experimental subjects were placed 3 h after 'sampling' for 4 min in a novel cage (right). (b) Transferring the home cage with the experimental subject in a new room shortly before 'sampling' (left) or transfer and removing the experimental subject from the cage and applying a brief (2–3 min) isoflurane anesthesia (right) failed to impair the memory performance at a retention interval of 24 h. Paired t-tests were performed for each 'choice' session separately. For more details see Figure 7.
Interference phenomena. Social memory is very susceptible to interference phenomena47, which can be monitored in both rats and mice and during short-term8,9 and long-term memory20 (Supplementary Fig. 2). Retroactive interference occurs when presenting a novel item during the retention interval and impairs the memory of the originally learned item (Supplementary Fig. 2b). Proactive interference occurs when a learning event in the past interferes with the acquisition of information about a novel item (Supplementary Fig. 2c). A recent extensive study investigated the two different types of interference for long-term social memory using the social discrimination procedure in mice20.
Materials
REAGENTS
-
Laboratory rats (Rattus norvegicus domestica) and mice (Mus musculus domestica; obtained, e.g., from Charles River or Harlan-Winkelmann) of different standard inbred and outbred strains. (see REAGENT SETUP)
Caution
Please adhere to all relevant institutional and governmental rules regarding the use of experimental animals (e.g., EEC recommendations for the care and use of laboratory animals (86/609/CEE)).
EQUIPMENT
-
Testing room: relatively small (∼10 m2), quiet, follows standards for animal housing rooms (air conditioning, controlled light/dark cycle, etc.), room luminosities between 135 and 250 lux
-
Tables with separate shelves 50 cm below the tables' surface (Fig. 4a)
Figure 4: Equipment setup. 
(a) Complete setup of the experimental room including a custom-made regular table (1.2 × 0.8 × 0.7 m) equipped with a board 0.5 m below the desktop. The cages for the experimental subjects (Es, 1–4) are arranged on the desktop and those for the corresponding juveniles (J1–J4) on the separate board below the desktop. (b) View of the experimenter into the mirror providing sight of the rear of the type III cage (arrows) to make it easier to observe the animals' behavior.
-
Polycarbonate type II cages (mice: 22 × 16 × 14 cm) or type III cages (rats: 37 × 21 × 15 cm) with fresh bedding for the experimental subjects
-
Polycarbonate type II cages (both species) for the stimulus animals
-
Computer with appropriate software
-
A chair for the observer
-
A mirror to facilitate behavioral observation in rats (Fig. 4b)
-
Edding 30 permanent marker (tip 1.5–3 mm, filled with neutral smelling water-based ink, green; Edding International) for labeling the juveniles
-
Disposable gloves for handling the animals
-
Volatile fraction cage (optional): see Box 1 and Figure 5
Figure 5: The volatile fraction cage. 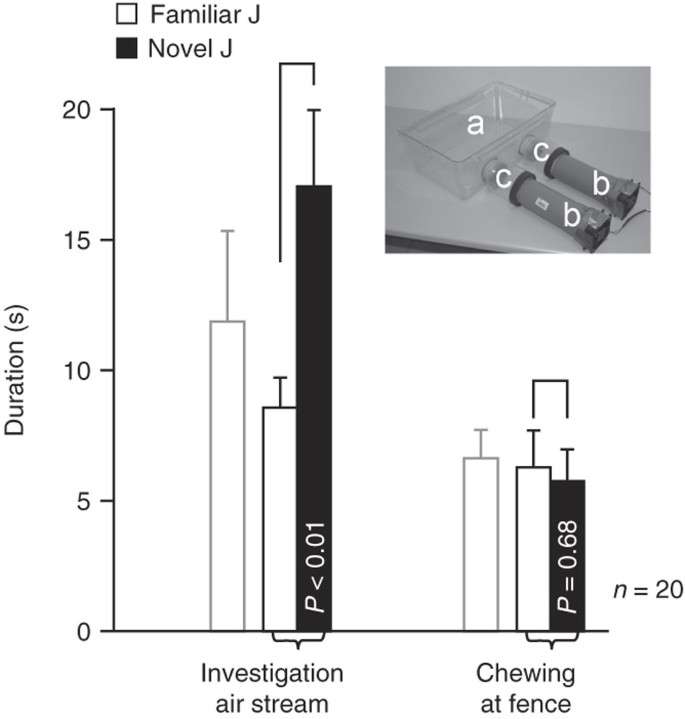
The inset shows a photograph of the volatile fraction cage. Experimental subjects are isolated in a transparent polycarbonate cage (39 × 24 × 16 cm, a) with two holes in one sidewall. To expose the experimental subjects to the volatile fraction of the juvenile's olfactory signature, a juvenile is put into the transparent plastic section (c) of the fan unit (b). The fan units allow the volatile components of the olfactory signature to diffuse into the experimental cage. The graph shows a representative example of the investigation duration of adult male mice toward the air stream and the duration of chewing at the fences separating the experimental subjects from the fan units. The experimental subjects spent significantly longer investigating the air stream containing the volatile fraction of the new juvenile than that of the familiar juvenile, whose odors' volatile fraction had been presented during 'sampling' 24 h before. In contrast, the nonspecific duration of chewing at the fences did not differ.



REAGENT SETUP
Experimental subjects
-
Use adult animals (approximately 8–16 weeks of age) of either species or sex.
Stimulus animals
-
Use group-housed (for each litter separately), conspecific juveniles (rats or mice, age between 15 and 35 d) of either sex. Note that it has been reported that adult ovariectomized females may also serve as stimulus animals12.
EQUIPMENT SETUP
Testing room
-
Along the walls in the testing room, regular tables serve as racks for the cages. Put the cages of the juvenile stimulus animals on the shelf below the tables' surfaces. To avoid confusion about the animals' assignments, label cages correctly or make sure that the cage of each stimulus animal exactly matches the above-placed cage of an experimental subject (Fig. 4a).
Procedure
Animal care
Timing 2 weeks
-
1
Acclimatize all animals to their groups and the housing conditions for 2 weeks.
-
2
Carefully check for bite wounds once per day, and, if necessary, observe the behavior of the animals in their home cage. Make sure to remove aggressive individuals (this is particularly important for studies of mice because they tend to be more aggressive than rats).
Separation of animals
Timing 2.5 h
-
3
Testing can be done at any point during the whole light-dark cycle. On the day of experiment, transfer the animals to the testing room and separate experimental subjects about 2 h before 'sampling' to allow them to become accustomed to the cage. Use gloves when handling the animals. At this time, separate and label half of the stimulus animals (with the Edding marker) at the head and/or at the base of the tail (Supplementary Fig. 3). Place the animals in their testing cages at the appropriate position in the room. Put only 4–6 food pellets in the cage rack to make it easy to remove all pellets just before testing. Put the water bottle in the cage rack.
Critical Step
Avoid any type of perfume or volatile odors (e.g., cigarette smoke) that may affect social investigation behavior.
Critical Step
Make sure that the experimental and stimulus animals are properly arranged and that the total duration needed to test subsequent experimental subjects is not longer than the exposure interval. For example, using a 30-min retention interval will allow a maximum of six experimental subjects to be tested sequentially (using a 'sampling' time of 4 min) before the second exposure ('choice') has to be started for the first animal (please take into account the time for inserting and removing the stimulus animals, i.e., start individual experimental subjects at 0, 5, 10 min and so on).
-
4
Switch on the computer and start the appropriate program and, if you wish, leave the room quietly until the testing starts.
'Sampling'
Timing 5 min per experimental subject
-
5
Put on gloves. Start the timer to ensure that the exposure interval is similar for all experimental subjects to be tested.
-
6
Remove bottles and food pellets from the cages of both the stimulus animal and the experimental subject to be tested.
-
7
Take the 'to be recognized' stimulus animal out of its home cage and place it in the cage of the corresponding experimental subject above (Fig. 6a).
Figure 6: Schematic diagram showing the sequence of juvenile placement into the experimental subjects' cages during 'sampling' and 'choice'. 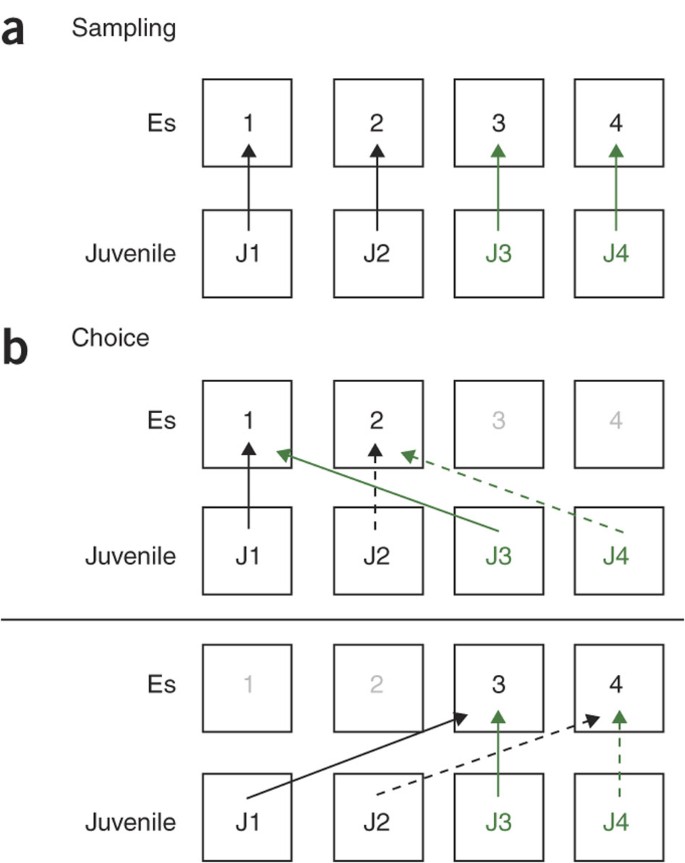
(a) 'Sampling'. The 'to-be-recognized' juvenile is placed into the cage of the corresponding experimental subject (Es; see Fig. 4a) just above (arrows). The representative protocol uses two unmarked (black J1 and J2) and two marked juveniles (green J3 and J4). (b) 'Choice'. The familiar juvenile (that had been presented during 'sampling') is placed, together with a novel juvenile, into the cage of the experimental subject. This setup allows a minimum of exposures for the juveniles and ensures that labeled and nonlabeled juveniles are always presented together. For simplicity, the 'choice' sequence is split into two separate panels. The upper panel shows juvenile use for experimental subjects 1 and 2, the lower panel shows the juvenile use for experimental subjects 3 and 4. Arrows with the same dash type indicate the simultaneous transfer of the juveniles to the cage of the given experimental subject. During 'sampling' and 'choice', the investigatory behavior and aggressive/sexual behavior are monitored by a trained observer using an appropriate computer program.
-
8
Start observation and behavioral scoring (Box 2). The exposure time is 4 min.
-
9
After the end of the observation period, remove the stimulus animal and return it to its home cage.
-
10
Repeat Steps 6–9 until all experimental subjects have been tested (Fig. 6a).


Retention interval
Timing variable
-
11
If the interval between 'sampling' and 'choice' is less than 1 h, do not put back the water bottle. If the interval is longer, return the bottles to the cages of the experimental subjects and stimulus animals. If the interval between 'sampling' and 'choice' is longer than 2 h, add 4–6 food pellets to each cage rack to ensure ad libitum feeding. Depending on the retention interval, you may now leave the room quietly until 'choice' starts.
'Choice'
Timing 5 min per experimental subject
-
12
If necessary, put on gloves, carefully remove the food pellets from the cage rack of the experimental subjects and remove the water bottles from the cages of the stimulus animal and of the experimental subject to be tested.
-
13
Take the previously encountered animal and a novel stimulus animal (Fig. 6b) out of their home cages, and place both in the cage of the corresponding experimental subject. Always be sure to use a labeled animal together with a nonlabeled stimulus animal to help the observer discriminate between the stimulus animals during testing.
-
14
Start observation and behavioral scoring (Box 2). The exposure time is 4 min.
-
15
After the end of the observation period, remove the stimulus animals and place them back in their individual home cages.
-
16
Repeat Steps 13–15 until all of the experimental subjects have been tested (Fig. 6b).
Post-test processing
Timing 1 h
-
17
Return all animals to their original groups; returning them to their original cages can facilitate habituation and reduce fighting.
Critical Step
Experimental subjects must be kept in their original groups for at least 1 d between any two tests. For the sake of stable cage cohorts, we usually run the social discrimination procedure with the same experimental subjects only once a week.
-
18
Save the data and/or close the computer program.
-
19
Analyze the data; parameters and their interpretation are outlined in Box 3 (see also Fig. 7). Please note that the representative group size for statistical analysis differs between rats and mice: in rats, groups should contain about 10 animals, whereas in mice, the groups should contain about 20 animals because of higher interindividual variability in behavioral performance.
Figure 7: Representative examples showing anticipated results obtained in the social discrimination procedure. 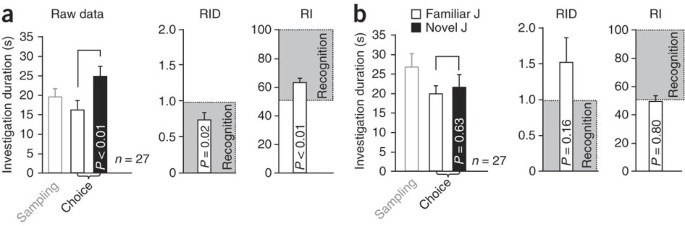
(a,b) Raw investigation durations (left), the ratio of investigation duration (RID, middle) and the recognition index (RI, right) based on the same raw data in a and b, respectively. Data were obtained from a representative sample of adult male mice; the retention interval was 24 h. In a, experimental subjects showed an intact recognition memory, indicated by a significant increase in the investigation duration toward the novel (novel J) versus the familiar juvenile (familiar J) during 'choice' (left panel, curly bracket; statistics: paired t test). This is mirrored by an RID significantly lower than 1 (1 = chance level) and an RI of significantly higher than 50 (50 = chance level), both calculated by one sample t-test. In b, experimental subjects failed to recognize the previously encountered juvenile, as indicated by statistically similar investigation durations for both juveniles during 'choice' (left panel; curly bracket; statistics: paired t test). This is mirrored by both the RID and RI, neither of which differ significantly from the chance level calculated as indicated above. Amnesia was produced by retroactive interference (presentation of a different juvenile during the retention interval).


Troubleshooting
Troubleshooting advice can be found in Table 2.
Timing
Steps 1 and 2, Animal care: ∼2 weeks
Steps 3 and 4, Separation of animals: ∼2.5 h
Steps 5–10, 'Sampling': ∼5 min per experimental subject
Step 11, Retention interval: variable; dependent on the experimental design and species
Steps 12–16, 'Choice': ∼5 min per experimental subject
Steps 17–19, Post-test processing: ∼1 h
Anticipated results
General parameters that are used to interpret the measurements obtained in the context of learning and memory are detailed in Box 2 and Box 3 illustrated in Figure 8. The primary parameter serving as an index for recognition is the ratio of the duration spent by the experimental subject during 'choice' (i.e., investigating the previously exposed stimulus animal) to that of the novel stimulus animal (Fig. 8b, left). As can be seen in Figure 8b (right), the number of investigation bouts may also be a suitable index measure if correspondingly analyzed. Figure 8c illustrates that aggressive and sexual behavior has a negligible role in untreated Wistar rats. In contrast, male (and female) C57BL/6JOlaHsd mice show more aggression and/or sexual behavior toward conspecific juveniles as stimulus animals during testing; this contributes 13–35% of the total number of behavioral bouts and the total duration of social behavior, respectively. Nevertheless, under untreated conditions in both standard mice and rat strains aggressive and/or sexual behavior does not interfere with the ability to measure the parameters used for analyzing social recognition memory.

(a) Representative event plot for olfactory investigation behavior monitored during 'choice' for a male mouse. The experimental subject investigates alternatively the (familiar J) and the novel juvenile (novel J) throughout the 4-min 'choice' session. (b) Raw investigation durations, number of investigation bouts and latencies to first investigate juveniles scored during 'choice' (curly bracket) at a retention interval of 24 h. Data were obtained from a representative sample of adult male mice. An intact recognition memory is indicated by a significantly longer investigation duration and by significantly more investigation bouts of the novel versus the familiar juvenile. In contrast, the latency to first investigate the juveniles failed to show a significant difference. Statistics was performed by paired t-test. (c) Differences in the frequency (and 'duration') of aggressive and sexual behavior during 'sampling' between rats and mice. Data are compiled from representative samples of male rats (white bars) and mice (black bars). For completeness, the inset also shows the 'duration' measured. The average investigation bout was too brief to allow for a reliable measurement; nevertheless, the data demonstrate that mice show more aggressive behavior than rats during social discrimination testing. Groups were compared by unpaired t-tests.
Unlike most memory tasks based on classical and instrumental conditioning or spatial navigation, the social discrimination test measures the presence or absence of recognition categorically, in the manner of a 'yes' or 'no' decision. However, more subtle differences in the memory performance can also be detected, for instance, as reflected by different memory retention intervals. A typical example is the aforementioned ability of female rats to recognize a conspecific 120 min after sampling, when social memory in male rats is already extinct26,31,33. This suggests a slightly better social memory performance in females within the constraints of short-term memory, as memory extinction after 180 min has also been reported in females31,33.
The social discrimination procedure enables the investigation of the impact of manipulations on the different 'stages' of memory. In particular, manipulations shortly before/during 'sampling' primarily affect acquisition, manipulations after 'sampling' affect memory consolidation and manipulations shortly before 'choice' affect memory retrieval. Treatments that interfere with recognition memory are expected to extinguish the difference between the investigation durations seen during 'choice' under standard conditions22,26,33. In contrast, a treatment that produces a markedly longer investigation duration of the novel versus the familiar stimulus animal during 'choice' is considered to have caused a memory performance–enhancing effect24,25,26.
Together, the findings listed above provide a framework for using the social discrimination task to select a detailed experimental design for testing the anticipated consequences of manipulations on the declarative memory performance of rodents.
References
Ebbinghaus, H. Über das Gedächtnis—Untersuchungen zur experimentellen Psychologie (Duncker & Humblot, Leipzig, 1885).
Schacter, D.L. Implicit memory: history and current status. J. Exp. Psychol.: Learning, Memory, and Cognition 13, 501–518 (1987).
Steckler, T., Drinkenburg, W.H., Sahgal, A. & Aggleton, J.P. Recognition memory in rats—I. Concepts and classification. Prog. Neurobiol. 54, 289–311 (1998).
Accolla, R. & Carleton, A. Internal body state influences topographical plasticity of sensory representations in the rat gustatory cortex. Proc. Natl. Acad. Sci. USA 105, 4010–4015 (2008).
McGaugh, J.L. Make mild moments memorable: add a little arousal. Trends Cogn. Sci. 10, 345–347 (2006).
Kamprath, K. & Wotjak, C.T. Nonassociative learning processes determine expression and extinction of conditioned fear in mice. Learn Mem. 11, 770–786 (2004).
Dantzer, R., Bluthe, R.M. & Kelley, K.W. Androgen-dependent vasopressinergic neurotransmission attenuates interleukin-1-induced sickness behavior. Brain Res. 557, 115–120 (1991).
Thor, D.H. & Holloway, W.R. Social memory of the male laboratory rat. J Comp. Physiol. Psychol. 96, 1000–1006 (1982).
Dantzer, R., Bluthe, R.M., Koob, G.F. & Le Moal, M. Modulation of social memory in male rats by neurohypophyseal peptides. Psychopharmacology (Berl.) 91, 363–368 (1987).
Engelmann, M., Wotjak, C.T. & Landgraf, R. Social discrimination procedure: an alternative method to investigate juvenile recognition abilities in rats. Physiol. Behav. 58, 315–321 (1995).
Reijmers, L.G., Leus, I.E., Burbach, J.P., Spruijt, B.M. & van Ree, J.M. Social memory in the rat: circadian variation and effect of circadian rhythm disruption. Physiol. Behav. 72, 305–309 (2001).
Sterlemann, V. et al. Chronic social stress during adolescence induces cognitive impairment in aged mice. Hippocampus 20, 540–549 (2010).
Choleris, E. et al. Involvement of estrogen receptor alpha, beta and oxytocin in social discrimination: a detailed behavioral analysis with knockout female mice. Genes Brain Behav. 5, 528–539 (2006).
Macbeth, A.H., Edds, J.S. & Young, W.S. III Housing conditions and stimulus females: a robust social discrimination task for studying male rodent social recognition. Nat. Protoc. 4, 1574–1581 (2009).
Bluthe, R.M. & Dantzer, R. Role of the vomeronasal system in vasopressinergic modulation of social recognition in rats. Brain Res. 604, 205–210 (1993).
Noack, J. et al. Different importance of the volatile and non-volatile fractions of an olfactory signature for individual social recognition in rats versus mice and short-term versus long-term memory. Neurobiol. Learn. Mem. 94, 568–575 (2010).
Popik, P., Vetulani, J., Bisaga, A. & van Ree, J.M. Recognition cue in the rat's social memory paradigm. J. Basic Clin. Physiol. Pharmacol. 2, 315–327 (1991).
Page, D.T., Kuti, O.J. & Sur, M. Computerized assessment of social approach behavior in mouse. Front Behav. Neurosci. 3, 48 (2009).
Insel, T.R. & Fernald, R.D. How the brain processes social information: searching for the social brain. Annu. Rev. Neurosci. 27, 697–722 (2004).
Engelmann, M. Competition between two memory traces for long-term recognition memory. Neurobiol. Learn. Mem. 91, 58–65 (2009).
Squires, A.S., Peddle, R., Milway, S.J. & Harley, C.W. Cytotoxic lesions of the hippocampus do not impair social recognition memory in socially housed rats. Neurobiol. Learn. Mem. 85, 95–101 (2006).
Richter, K., Wolf, G. & Engelmann, M. Social recognition memory requires two stages of protein synthesis in mice. Learn. Mem. 12, 407–413 (2005).
Wanisch, K., Wotjak, C.T. & Engelmann, M. Long-lasting second stage of recognition memory consolidation in mice. Behavioural. Brain Res. 186, 191–196 (2008).
Dluzen, D.E., Muraoka, S., Engelmann, M., Ebner, K. & Landgraf, R. Oxytocin induces preservation of social recognition in male rats by activating alpha-adrenoceptors of the olfactory bulb. Eur. J. Neurosci. 12, 760–766 (2000).
Dluzen, D.E., Muraoka, S., Engelmann, M. & Landgraf, R. The effects of infusion of arginine vasopressin, oxytocin, or their antagonists into the olfactory bulb upon social recognition responses in male rats. Peptides 19, 999–1005 (1998).
Landgraf, R. et al. V1 vasopressin receptor antisense oligodeoxynucleotide into septum reduces vasopressin binding, social discrimination abilities, and anxiety-related behavior in rats. J. Neurosci. 15, 4250–4258 (1995).
Liebsch, G., Landgraf, R., Engelmann, M., Lorscher, P. & Holsboer, F. Differential behavioural effects of chronic infusion of CRH 1 and CRH 2 receptor antisense oligonucleotides into the rat brain. J. Psychiatr. Res. 33, 153–163 (1999).
Tobin, V.A. et al. An intrinsic vasopressin system in the olfactory bulb is involved in social recognition. Nature 464, 413–417 (2010).
Jüch, M. et al. Congenital lack of nNOS impairs long-term social recognition memory and alters the olfactory bulb proteome. Neurobiol. Learn. Mem. 92, 469–484 (2009).
Kogan, J.H., Frankland, P.W. & Silva, A.J. Long-term memory underlying hippocampus-dependent social recognition in mice. Hippocampus 10, 47–56 (2000).
Bluthe, R.M. & Dantzer, R. Social recognition does not involve vasopressinergic neurotransmission in female rats. Brain Res. 535, 301–304 (1990).
Bluthe, R.M., Schoenen, J. & Dantzer, R. Androgen-dependent vasopressinergic neurons are involved in social recognition in rats. Brain Res. 519, 150–157 (1990).
Engelmann, M., Ebner, K., Wotjak, C.T. & Landgraf, R. Endogenous oxytocin is involved in short-term olfactory memory in female rats. Behav. Brain Res. 90, 89–94 (1998).
Engelmann, M. & Landgraf, R. Microdialysis administration of vasopressin into the septum improves social recognition in Brattleboro rats. Physiol. Behav. 55, 145–149 (1994).
Sanchez-Andrade, G. & Kendrick, K.M. Roles of alpha- and beta-estrogen receptors in mouse social recognition memory: effects of gender and the estrous cycle. Horm. Behav. 59, 114–122 (2011).
Terranova, J.P., Perio, A., Worms, P., Le Fur, G. & Soubrie, P. Social olfactory recognition in rodents: deterioration with age, cerebral ischaemia and septal lesion. Behav. Pharmacol. 5, 90–98 (1994).
Prediger, R.D., Batista, L.C. & Takahashi, R.N. Caffeine reverses age-related deficits in olfactory discrimination and social recognition memory in rats. Involvement of adenosine A1 and A2A receptors. Neurobiol. Aging 26, 957–964 (2005).
Bothe, G.W.M., Bolivar, V.J., Vedder, M.J. & Geistfeld, J.G. Behavioral differences among fourteen inbred mouse strains commonly used as disease models. Comparative Med. 55, 326–334 (2005).
Camp, M. et al. Impaired Pavlovian fear extinction is a common phenotype across genetic lineages of the 129 inbred mouse strain. Genes, Brain and Behav. 8, 744–752 (2009).
Cook, M.N., Bolivar, V.J., McFadyen, M.P. & Flaherty, L. Behavioral differences among 129 substrains: implications for knockout and transgenic mice. Behav. Neurosci. 116, 600–611 (2002).
Hefner, K. et al. Impaired fear extinction learning and cortico-amygdala circuit abnormalities in a common genetic mouse strain. J. Neurosci. 28, 8074–8085 (2008).
Owen, E.H., Logue, S.F., Rasmussen, D.L. & Wehner, J.M. Assessment of learning by the Morris water task and fear conditioning in inbred mouse strains and F1 hybrids: implications of genetic background for single gene mutations and quantitative trait loci analyses. Neuroscience 80, 1087–1099 (1997).
Simpson, E.M. et al. Genetic variation among 129 substrains and its importance for targeted mutagenesis in mice. Nat. Genet. 16, 19–27 (1997).
Thor, D.H. Testosterone and persistence of social investigation in laboratory rats. J. Comp. Physiol. Psychol. 94, 970–976 (1980).
Bluthe, R.M., Gheusi, G. & Dantzer, R. Gonadal steroids influence the involvement of arginine vasopressin in social recognition in mice. Psychoneuroendocrinology 18, 323–335 (1993).
Prediger, R.D., Fernandes, D. & Takahashi, R.N. Blockade of adenosine A2A receptors reverses short-term social memory impairments in spontaneously hypertensive rats. Behav. Brain Res. 159, 197–205 (2005).
Müller, G.E. & Pilzecker, A. Experimentelle Beiträge zur Lehre vom Gedächtnis. Zeitschrift für Psychologie und Physiologie der Sinnesorgane EB 1–300 (1900).
Burman, O.H. & Mendl, M. Short-term social memory in the laboratory rat: its susceptibility to disturbance. Appl. Anim. Behav. Sci. 67, 241–254 (2000).
Amano, K. et al. Estimation of the timing of human visual perception from magnetoencephalography. J. Neurosci. 26, 3981–3991 (2006).
Welker, W.I. Analysis of sniffing of the albino rat. Behaviour 22, 223–244 (1964).
Wesson, D.W., Donahou, T.N., Johnson, M.O. & Wachowiak, M. Sniffing behavior of mice during performance in odor-guided tasks. Chem. Senses 33, 581–596 (2008).
Kimchi, T., Xu, J. & Dulac, C. A functional circuit underlying male sexual behaviour in the female mouse brain. Nature 448, 1009–1014 (2007).
Perio, A. et al. Specific modulation of social memory in rats by cholinomimetic and nootropic drugs, by benzodiazepine inverse agonists, but not by psychostimulants. Psychopharmacology (Berl.) 97, 262–268 (1989).
Acknowledgements
We thank R. Murau and M. Radscheidt for expert support in behavioral testing. We also thank G. Leng, Edinburgh, for polishing the English. During the writing of this paper, the authors were supported by the Deutsche Forschungsgemeinschaft (EN 366/6-1 and EN 366/8-1).
Author information
Authors and Affiliations
Contributions
M.E. and J.N. designed the study, J.H. performed some of the behavioral experiments, and M.E. and J.N. wrote the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Fig. 1
No impact of female sex cycle on long-term social discrimination. The same group-housed female mice were tested for their social recognition abilities under estrous and non-estrous conditions at an retention interval of 24 h. The stage of the cycle was determined using microscopical analysis of vaginal smears taken ∼ 1 h after choice. Statistical analysis shows that experimental subjects successfully discriminated between the stimulus animals independently upon cyclus stage during choice. Statistics was performed using paired t-test. (JPG 329 kb)
Supplementary Fig. 2
Social discrimination is sensitive to interference phenomena. Examples showing schematically the experimental design (insets) and representative results obtained with same group-housed adult male mice as experimental subjects without (A) and with experimental induction of interference (B,C) for long-term social recognition memory. A, demonstrates intact recognition memory performance of these animals in the standard social discrimination protocol. B, Presentation of an interference juvenile (hatched) during the retention interval of 24h (6h after sampling) impairs recognition of the familiar juvenile. C, Presentation of an interference juvenile (hatched) 3h before sampling impairs recognition of the familiar juvenile at the retention interval of 21 h. Statistics was performed using paired t-test. Please note that the effect of interference is time-dependent. (JPG 533 kb)
Supplementary Fig. 3
Stimulus animal labelling. Schematic drawing showing the relative position and length of the marks used for labelling at the dorsal side of a juvenile. (JPG 165 kb)
Rights and permissions
About this article
Cite this article
Engelmann, M., Hädicke, J. & Noack, J. Testing declarative memory in laboratory rats and mice using the nonconditioned social discrimination procedure. Nat Protoc 6, 1152–1162 (2011). https://doi.org/10.1038/nprot.2011.353
Published:
Issue Date:
DOI: https://doi.org/10.1038/nprot.2011.353
This article is cited by
-
Detection, processing and reinforcement of social cues: regulation by the oxytocin system
Nature Reviews Neuroscience (2023)
-
A quantitative analysis of spontaneous alternation behaviors on a Y-maze reveals adverse effects of acute social isolation on spatial working memory
Scientific Reports (2023)
-
Dynamic and stable hippocampal representations of social identity and reward expectation support associative social memory in male mice
Nature Communications (2023)
-
The extracellular matrix and perineuronal nets in memory
Molecular Psychiatry (2022)
-
Top-down acetylcholine signaling via olfactory bulb vasopressin cells contributes to social discrimination in rats
Communications Biology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.

{kind=link}
{kind=link}
{kind=link}