
Abstract
Elevated atmospheric CO2 concentration ([CO2]) can stimulate plant growth through enhanced photosynthetic rate. However, plant C, N and P ratios in response to elevated [CO2] combined with canopy warming in rice-winter wheat rotation system remain largely unknown. Here we investigated the impacts of elevated [CO2] and warming on plant nutrient ratios under open-air conditions. Four treatments including the ambient condition (CK), elevated [CO2] (500 ppm, CE), canopy warming (+2 °C, WA), and the combination of elevated [CO2] and warming (CW) were used to investigate the responses of plant C, N and P ratios in a rice-winter wheat rotation system in southeast China. Results showed that elevated [CO2] increased C:N ratio in whole plant by 8.4–14.3% for both crops, and increased C:P ratio by 11.3% for rice. The changes in ratio were due to an increase in C concentration by 0.8–1.2% and a reduction in N concentration by 7.4–10.7% for both crops, and a reduction in P concentration by 10.0% for rice. Warming increased N allocation in rice leaf and N concentration by 12.4% for rice, resulting in increases in the ratios of N to C and P by 11.9% and 9.7% in rice, but not in wheat. However, CW had no effect on plant C:N ratio in rice, indicating the positive effect of elevated [CO2] could offset the negative impact of warming on C:N ratio. By contrast, CW significantly decreased plant C:P and N:P ratios by 16% due to the increase in P allocation in stem for wheat. These results suggest that impacts of climate change on plant nutrient balance occur through interactions between the effects of climate change on nutrient uptake and allocation, which is important for food quality and productivity under global climate change.
Similar content being viewed by others


Introduction
Global atmospheric carbon dioxide concentration ([CO2]) has increased rapidly due to the ongoing anthropogenic activities since the industrial revolution1. It is a common belief that elevated [CO2] stimulates terrestrial plant growth by enhancing CO2 fixation rate, known as the [CO2] fertilization effect2,3,4. Elevated [CO2] also benefits plant growth in drought regions by promoting water use efficiency via reduction in stomatal conductance5,6. However, plant growth promotion might be constrained by the nutrient deficiency under elevated [CO2], as the plants demand more nutrients for growth under elevated [CO2]7. Previous studies suggested that future plants would be exposed to a global nutrient imbalance under elevated [CO2]8,9,10,11, with lower N concentration10 or increased C:N and C:P ratios in plant functional organs9. However, the response of C:N or C:P ratio to elevated [CO2] showed large variation across various ecosystems11. This might be resulted from the different nutrient management regimes across these systems. In forest or grassland ecosystems, progressive occurrence of nutrient limitation under elevated [CO2] may occur as fertilization are seldom performed7. In contrast, in agroecosystems nutrient limitation may not occur under elevated [CO2] due to sufficient nutrient supply from fertilizers. Nevertheless, some studies reported that elevated [CO2] increased C:N ratio12,13. So far, it remains unclear how elevated [CO2] impact the direction and magnitude of nutrient uptake and their ratios in agroecosystems.
It is increasingly important to recognize that the responses of plants to elevated [CO2] are associated with other factors that change terrestrial ecological processes, in particular global warming. Global average surface temperature is estimated to rise by 1.1–6.4 °C at the end of 21st century14. Unlike elevated [CO2], global warming can either decrease plant productivity by shortening plant growing period and reducing panicle number in warm regions15,16,17,18 or stimulate plant grown in cold regions19,20. Under a combination of elevated [CO2] and warming, the overall nutrient uptake and their ratio to carbon in plants can also be changed. Some studies have reported that warming increase nutrient concentrations in plants21,22. However, Yuan and Chen11 reported that warming alone had no effect on plant N concentration but decreased plant P concentration across a range of plant communities. In comparison, the combination of elevated [CO2] and warming had a neutral effect on N and P concentrations and N:P ratio in plants11,23.
In addition to nutrient concentrations and their ratios in the whole plants, it is increasingly important to understand the nutrient allocation across different plant functional organs. In some studies, nutrient concentrations of the whole plant were not affected by elevated [CO2] or warming. However, being unresponsive in the whole plant does not necessarily mean that the nutrient concentrations in a specific functional organ is not affected. Inconsistent responses of plant nutrient concentrations across different functional organs have been reported13,22,24,25. For instance, Cheng et al. found that elevated night (from 20:00 until 04:00) temperature by ca 10 °C had no effect on whole rice plant C or N concentration, but increased N concentration in the living leaf and reduced its allocation to the ear13. Results from these studies were mainly dependent on plant types and experimental conditions. To date, the effect of elevated [CO2] and canopy warming on plant nutrient (C, N and P) ratios in a rice-wheat rotation system is not well understood.
The main objective of this study was to examine the changes in plant nutrient (C, N and P) uptake and their ratios in different plant functional organs in a summer rice-winter wheat rotation system under simulated climate change conditions. We hypothesized that elevated [CO2] had a positive effect on C to N and C to P ratios due to dilution effect; warming would increase nutrient concentrations, and alter plant C, N and P ratios under elevated [CO2] and warming. We aim to provide knowledge for improving food quality and nutrient management in agriculture under future climate change.
Results
C, N and P concentrations in the whole plant
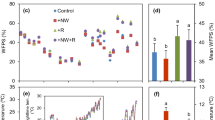
Average cross three crop growth stages, elevated [CO2] increased C concentration by 1.2% and 0.8% for rice (p = 0.001) and wheat (p = 0.061), respectively; whereas warming decreased C concentration by 0.8% for rice (p < 0.05) (Fig. 1). A significant interaction between [CO2] and warming was observed for C concentration in rice (p < 0.05). Elevated [CO2] decreased N concentration by 10.7% and 7.4% for rice and wheat (p < 0.001); and warming increased that by 12.4% and 10.5% for rice and wheat (p < 0.001), respectively. Elevated [CO2] decreased P concentration by 10.0% in rice (p < 0.05), but not in wheat (p = 0.156). In contrast, warming increased P concentration by 14.8% in wheat (p < 0.001), but not in rice (p = 0.186).

Changes in C, N and P concentrations in whole plant for rice (gray histogram) and wheat (black histogram) across three growth stages under simulated climate change conditions. Different letters indicate significant differences between treatments at p < 0.05.
The responses of C concentration to elevated [CO2] in different organs were similar to the whole plant (Tables S1, S2). For wheat, all treatments increased C concentration in stem and leaf at the ripening stage. However, the responses of N concentration to elevated [CO2] and warming varied with plant organs. Elevated [CO2] decreased N concentration by 21.5% and 7.1% in leaf for rice and wheat. Warming increased N concentration by 21.8% in leaf for rice, but decreased that for wheat. However, warming increased N concentration by 6.1% and 6.7% for rice (p < 0.01) and wheat (p = 0.075), respectively. The responses of P concentration to treatments also varied with organs. Elevated [CO2] increased P concentration by 11.8% and 10.5% in panicle/spike for rice (p = 0.010) and wheat (p = 0.056), while decreased that in stem and leaf of rice, and had no effect for wheat. However, warming increased P concentration in rice panicle and in wheat stem, but had no significant effect in other organs.
Plant C, N and P ratios
Plant C, N and P ratios in the whole plant exerted some remarkable changes under the treatments for both crops (Tables S3, S4). Generally, elevated [CO2] increased C:N ratio by 14.3% and 8.4% (p < 0.001), while warming reduced C:N ratio by 11.9% and 11.4% for rice and wheat (p < 0.001), respectively (Tables S3, S4). However, CW had no effect on C:N ratio in the whole plant for both crops (Fig. 2). Elevated [CO2] increased C:P ratio by 11.3% in the whole rice (p = 0.015), while warming had no effect on that. In contrast, warming decreased C:P ratio by 14.4% in the whole wheat, but the ratio was not affected by elevated [CO2]. Warming increased the N:P ratio by 9.7% in the whole rice (p < 0.05). Elevated [CO2] decreased the N:P ratio by 10.9% in the whole wheat (p < 0.01). Interaction effects between crop growth stages and treatments were observed in this study (Tables S3, S4). For example, elevated [CO2] significantly increased C:P ratio at the elongation and heading stages of rice, but not at the ripening stage.

Changes in C:N, C:P and N:P ratios in whole plant for rice (gray histogram) and wheat (black histogram) across three growth stages under simulated climate change conditions. Different letters indicate significant differences between treatments at p < 0.05.
The responses of C, N and P ratios across plant stem, leaf and panicle/spike varied between rice and wheat (Tables S3 and S4). For rice, leaf and panicle were more sensitive to elevated [CO2] and warming than stem; while stem was more responsive than leaf and spike for wheat. More specifically, elevated [CO2] increased the C:N ratio by 29.3% in rice leaf while warming alone decreased that by 20.1% (p < 0.001) (Table S3). For rice panicle, elevated [CO2] and warming generally decreased the C:N, C:P and N:P ratios. For wheat, elevated [CO2] had no significant effect on C, N and P ratios in different organs. However, warming decreased C:N and N:P ratios in stem, but increased C:N and C:P ratios in leaf for wheat.
C, N and P allocations across organs
At the elongation stage, elevated [CO2], warming or their combination had no effect on C allocation across leaf or stem (Fig. 3a,b). However, at the heading and ripening stages, elevated [CO2] and warming significantly affected C allocation. Elevated [CO2] reduced the C allocation from stem to panicle/spike for both crops, but had no significant effect on leaf. At the ripening stage, warming alone generally decreased C allocation in panicle/spike for rice, and increased it for wheat. Additionally, warming increased the allocation of C in rice leaf. CW increased C allocation in rice panicle.

Effects of elevated [CO2] and warming on the amount of C (above panel), N (middle panel) and P (below panel) distribution in stem, leaf and panicle (pike) for rice (a,c,e) and wheat (b,d,f). Different letters indicate significant differences between treatments at p < 0.05.
The responses of N allocation to elevated [CO2] and warming differed between rice and wheat (Fig. 3c,d). Elevated [CO2] decreased N allocation in panicle at the heading and ripening stages in rice, but not in wheat. Warming increased N allocation in rice stem and leaf at the ripening stage. For wheat, warming decreased N allocation in leaf, but increased it in panicle/spike at the ripening stage. CW decreased N allocation in the panicle of rice, but increased it in the panicle/spike of wheat.
Treatment effects on P allocation varied across growth stages for both crops (Fig. 3e,f). For rice, none of the treatments had significant effect on P allocation except for a significant increase in plant panicle under CW at the heading stage. For wheat, CW decreased P allocation in leaf at the elongation and ripening stages (Fig. 3e). Elevated [CO2] decreased P allocation in leaf but increased P allocation in spike at the heading stage. Warming alone increased P allocation in spike but decreased it in leaf and stem at the ripening stage.
Relation between C and nutrient (N and P)
The N and P accumulation were positively correlated with C accumulation for both crops (Fig. 4). Moreover, a negative correlation was also found between the C concentration and nutrient concentrations in stems for both crops (p < 0.05, Table 1).

Relationships between C accumulation and N accumulation (a), P accumulation (b) for rice (black regressions lines) and wheat (gray regressions lines) during elongation (white), heading (gray) and ripening (black) stages under simulated climate change conditions.
Discussion
Crop biomass and yield response and accordingly water and nutrient use efficiency, an issue of food supply and nutrition, had been already addressed in previous studies6,16,26,27,28. Our previous study shown that biomass ranged from 990 g m−2 to 1410 g m−2 for wheat, and from 1337 g m−2 to 1789 g m−2 for rice across the treatments16. Rice biomass was significantly greater than wheat. Elevated [CO2] increased crop biomass by 17.6% for both crops, while warming decreased biomass by 17.2% for wheat and 12.1% for rice. There was no significant effect between combined treatment and ambient condition. A similar trend was also observed in grain yield for both crops. However, the present study addresses how elevated [CO2] and warming impact on nutrients uptake and their ratios in different plant functional organs, an issue related to crop productivity and food nutrition value.
Effect of elevated [CO2] on plant C, N and P ratios
Our findings partially supported the hypothesis that elevated [CO2] increased C concentration, but reduced N and P concentrations for both crops. An increase in C:N and C:P ratios (except for wheat) was accordingly observed under elevated [CO2]. The decrease in N and P concentrations under elevated [CO2] may result from a dilution effect due to biomass increase (13.5–17.7%) under elevated [CO2] for both crops16. There was a negative correlation between C concentration and nutrient (N and P) concentrations in stem (Table 1). This is in agreement with previous studies9,11,29 conducted across various ecosystems. However, Yuan and Chen11 found that elevated [CO2] had no effect on C:N and C:P ratios in deciduous and evergreen woody angiosperms. This indicated that the responses of C:N and C:P ratios to elevated [CO2] varied greatly among plants.
To date, few experiments have tested the response of nutrient ratios to elevated [CO2] across different plant functional organs. We expected that different plant organs have the same responses to elevated [CO2]. However, our results showed that elevated [CO2] induced greater decrease in nutrient concentration in leaf than in stem and panicle/spike (Tables S1, S2). Elevated [CO2] generally increased leaf C:N and C:P ratios but had no effect on stem and panicle/spike. This is due to the different nutrient translocation patterns with crop growth. Therefore, elevated [CO2] induced plant nutrient imbalance, particularly C, N and P balance under future climates11.
Effect of warming on plant C, N and P ratios
Our results showed that warming alone decreased C:N ratio of rice because of the increased N concentration in rice and unchanged C concentration. Cheng et al. also observed that elevated night temperature had no effect on C concentration in rice despite an increase in plant respiration13. Additionally, An et al. reported that warming enhanced plant N concentration by 29.8–32.7% in a grassland ecosystem30. The change in N concentration could be ascribed to increased leaf transpiration rate under warming condition21. Warming increase plant leaf transpiration rate, leading to higher water requirement, which drives nutrient translocation from belowground to aboveground31,32. This is further confirmed by our observation that warming stimulated N transport from belowground. These results imply that global warming would exert a stronger effect on N uptake than on C assimilation, resulting in an imbalance in plant C and N content.
Additionally, warming alone had no effect on P concentration in the whole plant in this study (Fig. 1c), which led to an increase in N:P ratio in rice (Fig. 2). This suggests differential responses of N and P concentrations to warming alone. This might be explained by three possible reasons. Firstly, we inferred that the demand of N was more than that of P, as N is an important organ of organic compounds (e.g. amino acids, amides, proteins, nucleic acids, nucleotides, coenzymes, chlorophyll) for plant metabolism; whereas P is a component of sugar phosphates, which is less than N in plant functional organs33. Secondly, high temperature may have kinetic effects on the photosynthetic and respiration rate34,35, which requires more N input. Thirdly, our previous study has reported that warming had different effects on soil micronutrient availability through changes in soil environmental conditions (e.g. soil pH, moisture and microbial biomass)32. The increase of N:P ratio can be attributed to increases in soil nitrification rate and net N mineralization36, but reduction in soil P availability37. Therefore, warming increased N uptake from below ground, and influenced nutrient ratios in plant. As mentioned above, increment of N uptake in rice was higher than that of P. This is consistent with the study conducted by Reich and Oleksyn38, who also found that the N:P ratio increased with increment of air temperature.
Interestingly, different from rice, warming did not alter ratios of C, N and P in whole wheat (Fig. 2). Our results here demonstrated that warming did not alter N allocation and concentration in leaf for wheat, while a significant increase in N allocation to leaf was observed in rice (Fig. 3 and Tables S1, S2). Winter wheat was more sensitive than rice to warming17,39, resulting in limitation in wheat growth and N uptake under warming conditions. Furthermore, the N concentration increment of 13.4% in rice was higher than wheat (6.7%) under warming. This may be attributed to different N fertilizer input to rice (280.5 kg N ha−1) vs wheat (112.5 kg N ha−1). Therefore, the response in nutrient ratios to global warming would depend on crop types and agronomic management.
Effect of elevated [CO2] and warming on plant C, N and P ratios
Numerous studies have reported that elevated [CO2] altered nutrient balance by increasing carbohydrate production40,41,42,43, but few studies have considered the combined effects of elevated [CO2] and warming8,13,24. Our results revealed that the responses of plant nutrient ratios to a combination of elevated [CO2] and warming were remarkably differed from that to elevated [CO2] or warming alone (Fig. 2), because of the offset effects between elevated [CO2] and warming32,44. For example, C:N ratio was significantly increased by elevated [CO2] alone, decreased by warming alone (Fig. 2a), but was unaffected under the combined treatment for either crop. This indicates that the positive effect of elevated [CO2] on C:N ratio compensate for the negative impact of warming. This is similar to the changes in crop productivity under the combined effects of elevated [CO2] and warming. This indicates that the effects of elevated [CO2] offset the impacts of warming on crop growing and nutrient uptake16,17. Our study demonstrated that the combination of elevated [CO2] and warming had no effect on C or N concentration, but remarkably increased P concentration in wheat (Fig. 1). This was attributed to the responses of nutrient uptake and allocation to the combined treatment of elevated [CO2] and warming, which significantly increased P allocation and concentration in stem for wheat (Fig. 3f and Tables S1, S2). This was in agreement with Bhattacharyya et al.’s study, which showed that combined elevated [CO2] and warming significantly increased crop P uptake45. This is due to the increase in organic acid from root exudate and P mineralization under the combination of elevated [CO2] and warming, which solubilized P in soil42,45. This would lead to an imbalance of P with other elements in winter wheat field. Our previous study has found that the rice biomass was higher than wheat16, resulting in more P demand for rice. The mechanisms of nutrient cycling under combined effects of elevated [CO2] and warming may be more complex than elevated [CO2] or warming alone. This information is important for the sustainability of nutrient availability in agroecosystems under future climate change.
Conclusions
This study demonstrated that elevated [CO2] or warming alone significantly affected plant nutrient ratios in an agroecosystem, which varied with plant types and functional organs. Averaged across three key growth stages, elevated [CO2] increased C:N ratio for both crops mainly by reducing N concentration, whereas warming decreased C:N ratio while increased N:P ratio in rice due to enhanced N uptake and allocation of N in leaf. The combination of elevated [CO2] and warming had no effect on C:N ratio, but decreased C:P and N:P ratios in wheat. This was attributed to the increase in P allocation in wheat stem. The responses of nutrient uptake and ratios under combined elevated [CO2] and warming were different from that under elevated [CO2] or warming alone. This suggests that a offset effect exists between elevated [CO2] and warming. Therefore, the impact of climate change (elevated [CO2] and warming) on crop nutrient dynamics would be better predicted by a combination of these two factors rather than elevated [CO2] or warming alone.
Materials and Methods
Site description
This study was conducted in a field experiment station, where simulated elevated atmospheric [CO2], warming and their combination were performed to investigate the effect of climate change on an agroecosystem. The experimental site was established in 2010 and located at Kangbo village (31°30′N, 120°33′E), Guli Township, Changshu Municipality in Jiangsu Province, China. This area has a subtropical monsoon climate with a mean annual precipitation of 1100–1200 mm and annual average temperature of 16 °C over the last decade. The study area was a typical paddy field in the Taihu Lake region. The soil was a Gleyic Stagnic Anthrosol formed on clayey lacustrine deposit with a loamy texture (34% sand, 39% silt and 26% clay). The topsoil (0–20 cm) had 19.4 g kg−1 total C, 1.3 g kg−1 total N, 0.9 g kg−1 total P and a soil pH (H2O) of 7.027.
Experimental design
The experimental system was constructed under the state project of “Climate Change Impacts on Crop Production and Mitigation”, developed and managed by the Institute of Resource, Ecosystem and Environment of Agriculture (IREEA), Nanjing Agricultural University. The operational procedures of the facility were described in the work by Wang et al.16. In brief, the facility was designed to investigate two factors of climatic change, including elevated [CO2] of up to 500 ppm (CE), warming of canopy air by 2 °C with infrared heater over the crop canopy (WA), and a combination of these two treatments (CW), with ambient [CO2] without warming being the control (CK). Each treatment was deployed in an octagonal ring with a diameter of 8 m (area of ca 50 m2) and with three replications, totalling 12 rings. For elevated [CO2] treatments, pure CO2 gas via a liquid tank was injected into the ring plot with perforated pipes surrounding the ring. CO2 gas release was automatically manipulated based on ambient [CO2] and wind direction and speed. A total of 17 CO2 gas monitoring points were evenly distributed in each ring to determine the spatial variation of atmospheric [CO2]. Canopy air warming was performed with infrared heaters, hanging over the ring plot. A total of 12 infrared heaters (IR) (2000 W, 240 V, 1.65 m long × 0.14 m wide; HS-2420, Kalglo Electronics Co., Inc., Bethlehem, PA, USA) were equipped for each ring plot. The IR lamps produced invisible radiation to elevate the canopy air temperature. Both elevated [CO2] and canopy warming occurred throughout the crop growing period (rice, June-November; wheat, November-May). All the rings were separated by the adjacent open fields with about 28 m apart to avoid any treatment cross-contamination. The average [CO2] were 514 ± 55 ppm in rice season of 2012, and 505 ± 26 ppm in wheat season of 2012–2013 under the elevated [CO2] treatments, while the increment of canopy air temperature was a mean of 1.8 ± 0.6 °C in rice season of 2012, and 1.5 ± 0.7 °C in wheat season of 2012–2013 in the warming treatments, respectively. The performance and maintenance of these treatments were carefully managed throughout the crop growing periods.
Agronomic management
Rice (Oryza sativa L. cv. Changyou No. 5) seedlings were transplanted on 15th June 2012 and harvested on 28th October 2012. Winter wheat (Triticum aestivum L. cv. Yangmai No.14) was sown at a density of 250 seedlings m−2 with a row spacing of 20 cm on 23rd November 2012, and harvested on 2nd June 2013. Field management, including weed and pest control, fertilization and irrigation, was carried out following the local farmers’ practice. Fertilizers were applied four times during rice season and three times during wheat season. Urea (46% N) was applied as basal fertilizers at a rate of 188 kg ha−1, and as topdressing fertilizer at a rate of 150 kg ha−1 once in the wheat seasons and twice in the rice seasons. Compound fertilizer (N: P2O5: K2O ratio, 15:15:15) was applied at 375 kg ha−1 as topdressing after the heading stage for both crops. The total amount of N, P and K was applied at 212, 25 and 47 kg ha−1 in wheat seasons, and at 281, 25 and 47 kg ha−1 in rice seasons, respectively. The basal fertilizer was broadcast and incorporated into the soil by ploughing to the depth of plow layer. Paddy rice was irrigated with the regimes of continuous flooding, with two periods of drainage in mid-season.
Plant sampling and analysis
Plant samples were collected during the stem elongation, heading and ripening stages of rice in 2012 and of wheat in 2012–2013. The samples were separated into leaf, stem and ear. These samples were then oven-dried at 105 °C for 30 minutes and further oven-dried at 70 °C until constant weight. These oven-dried samples were then ground to yield a fine powder of <0.25 mm. Carbon concentration was measured with a CNS Macro Elemental Analyzer (Elementar, Germany). The plant leaf, stem and ear samples were pretreated with H2SO4-H2O2, dry-ashed at 550 °C for 4 h, and then the N concentration was determined with the Kjeldahl digestion method. Meanwhile, the P concentration (of acid digests) was analyzed by spectrophotometer (TU-1810, Beijing Purkinje General Instrument Co., Ltd., China).
Statistical analysis
Statistical analyses were carried out in SPSS22.0 (IBM SPSS Statistics for Windows, Version 22.0. Armonk, NY, USA). We used a general linear mixed model (GLM) to test the main effects of elevated [CO2], warming and growth stage (main factor), and their interaction on nutrients uptake and their ratios. A post hoc test was followed if any treatment effect was significant (p < 0.05). All data were presented as mean plus or minus standard deviations.
References
Houghton, J. T. et al. Climate change 2001: the scientific basis: contribution of Working Group I to the third assessment report of the Intergovernmental Panel on Climate Change. (Cambridge University Press 2001).
Ainsworth, E. A. Rice production in a changing climate: a meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration. Global Change Biology 14, 1642–1650, https://doi.org/10.1111/j.1365-2486.2008.01594.x (2008).
Cheng, W., Sakai, H., Yagi, K. & Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agricultural and Forest Meteorology 149, 51–58, https://doi.org/10.1016/j.agrformet.2008.07.006 (2009).
Yang, L. et al. Seasonal changes in the effects of free-air CO2 enrichment (FACE) on growth, morphology and physiology of rice root at three levels of nitrogen fertilization. Global Change Biology 14, 1844–1853, https://doi.org/10.1111/j.1365-2486.2008.01624.x (2008).
Xu, Z., Jiang, Y., Jia, B. & Zhou, G. Elevated-CO2 Response of Stomata and Its Dependence on Environmental Factors. Frontiers in Plant Science 7, https://doi.org/10.3389/fpls.2016.00657 (2016).
Wang, J. et al. Winter wheat water requirement and utilization efficiency under simulated climate change conditions: A Penman-Monteith model evaluation. Agricultural Water Management 197, 100–109, https://doi.org/10.1016/j.agwat.2017.11.015 (2018).
Luo, Y. et al. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 54, 731–739 (2004).
Gifford, R. M., Barrett, D. J. & Lutze, J. L. The effects of elevated [CO2] on the C: N and C: P mass ratios of plant tissues. Plant and Soil 224, 1–14 (2000).
Sardans, J., Rivas-Ubach, A. & Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol 14, 33–47, https://doi.org/10.1016/j.ppees.2011.08.002 (2012).
Cotrufo, M. F., Ineson, P. & Scott, A. Elevated CO2 reduces the nitrogen concentration of plant tissues. Global Change Biol 4, 43–54, https://doi.org/10.1046/j.1365-2486.1998.00101.x (1998).
Yuan, Z. Y. & Chen, H. Y. H. Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes. Nature Climate Change 5, 465–469, https://doi.org/10.1038/nclimate2549 (2015).
Liao, J., Hou, Z. & Wang, G. Effects of elevated CO2 and drought on chemical composition and decomposition of spring wheat (Triticum aestivum). Funct. Plant Biol 29, 891–897, https://doi.org/10.1071/PP01168 (2002).
Cheng, W., Sakai, H., Yagi, K. & Hasegawa, T. Combined effects of elevated [CO2] and high night temperature on carbon assimilation, nitrogen absorption, and the allocations of C and N by rice (Oryza sativa L.). Agricultural and Forest Meteorology 150, 1174–1181 (2010).
IPCC. Climate Change 2013: The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. (Cambridge University Press 2013).
Li, X., Jiang, D. & Liu, F. Soil warming enhances the hidden shift of elemental stoichiometry by elevated CO2 in wheat. Scientific Reports 6, 23313, https://doi.org/10.1038/srep23313 (2016).
Wang, J. et al. Size and variability of crop productivity both impacted by CO2 enrichment and warming—A case study of 4 year field experiment in a Chinese paddy. Agriculture, Ecosystems &. Environment 221, 40–49, https://doi.org/10.1016/j.agee.2016.01.028 (2016).
Ruiz-Vera, U. M. et al. Global warming can negate the expected CO2 stimulation in photosynthesis and productivity for soybean grown in the Midwestern United States. Plant Physiology 162, 410–423 (2013).
Matsui, T., Namuco, O. S., Ziska, L. H. & Horie, T. Effects of high temperature and CO2 concentration on spikelet sterility in indica rice. Field Crop. Res 51, 213–219 (1997).
Blumenthal, D. M., Kray, J. A., Ortmans, W., Ziska, L. H. & Pendall, E. Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland. Global Change Biol 22, 3026–3038, https://doi.org/10.1111/gcb.13278 (2016).
Usui, Y. et al. Rice grain yield and quality responses to free air CO2 enrichment combined with soil and water warming. Global Change Biology 22, 1256–1270 (2016).
Jauregui, I. et al. Nitrogen assimilation and transpiration: key processes conditioning responsiveness of wheat to elevated [CO2] and temperature. Physiologia plantarum 155, 338–354 (2015).
Kim, H.-Y. et al. Dry matter and nitrogen accumulation and partitioning in rice (Oryza sativa L.) exposed to experimental warming with elevated CO2. Plant and Soil 342, 59–71 (2011).
Van Groenigen, K. J., Qi, X., Osenberg, C. W., Luo, Y. & Hungate, B. A. Faster decomposition under increased atmospheric CO2 limits soil carbon storage. Science 344, 508–509 (2014).
Roy, K. S., Bhattacharyya, P., Neogi, S., Rao, K. S. & Adhya, T. K. Combined effect of elevated CO2 and temperature on dry matter production, net assimilation rate, C and N allocations in tropical rice (Oryza sativa L.). Field Crops Research 139, 71–79, https://doi.org/10.1016/j.fcr.2012.10.011 (2012).
Zhang, G. et al. The effects of free-air CO2 enrichment (FACE) on carbon and nitrogen accumulation in grains of rice (Oryza sativa L.). J. Exp. Bot 64, 3179–3188, https://doi.org/10.1093/jxb/ert154 (2013).
Wang, J. et al. Changes in nutrient uptake and utilization by rice under simulated climate change conditions: A 2-year experiment in a paddy field. Agricultural and Forest Meteorology 250–251, 202–208, https://doi.org/10.1016/j.agrformet.2017.12.254 (2018).
Wang, J. et al. Changes in micronutrient availability and plant uptake under simulated climate change in winter wheat field. Journal of Soils and Sediments 16, 2666–2675 (2016).
Wang, J. et al. Changes in grain protein and amino acids composition of wheat and rice under short term increased [CO2] and temperature of canopy air in a paddy from East China. New Phytologist 0, https://doi.org/10.1111/nph.15661 (2019).
Huang, W., Houlton, B. Z., Marklein, A. R., Liu, J. & Zhou, G. Plant stoichiometric responses to elevated CO2 vary with nitrogen and phosphorus inputs: Evidence from a global-scale meta-analysis. Sci Rep-UK 5, 18225 (2015).
An, Y. et al. Plant nitrogen concentration, use efficiency, and contents in a tallgrass prairie ecosystem under experimental warming. Global Change Biology 11, 1733–1744 (2005).
Trenberth, K. E., Fasullo, J. & Smith, L. Trends and variability in column-integrated atmospheric water vapor. Climate Dynamics 24, 741–758, https://doi.org/10.1007/s00382-005-0017-4 (2005).
Fernando, N. et al. Intra-specific variation of wheat grain quality in response to elevated [CO2] at two sowing times under rain-fed and irrigation treatments. J. Cereal Sci 59, 137–144 (2014).
Taiz, L. & Zeiger, E. Pant Physiology (4ed) (Chapter 5). 67–86 (Sinauer Associates, Inc 2006).
Gunderson, C. A., Norby, R. J. & Wullschleger, S. D. Acclimation of photosynthesis and respiration to simulated climatic warming in northern and southern populations of Acer saccharum: laboratory and field evidence. Tree Physiol 20, 87–96, https://doi.org/10.1093/treephys/20.2.87 (2000).
Atkin, O. Thermal acclimation and the dynamic response of plant respiration to temperature. Trends Plant Sci 8, 343–351, https://doi.org/10.1016/s1360-1385(03)00136-5 (2003).
Liu, Y. et al. Short-term response of nitrifier communities and potential nitrification activity to elevated CO2 and temperature interaction in a Chinese paddy field. Appl. Soil Ecol 96, 88–98, https://doi.org/10.1016/j.apsoil.2015.06.006 (2015).
Dijkstra, F. A. et al. Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland. The New phytologist 196, 807–815, https://doi.org/10.1111/j.1469-8137.2012.04349.x (2012).
Reich, P. B. & Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 101, 11001–11006, https://doi.org/10.1073/pnas.0403588101 (2004).
Ray, D. K., Gerber, J. S., MacDonald, G. K. & West, P. C. Climate variation explains a third of global crop yield variability. Nature communications 6, 5989, https://doi.org/10.1038/ncomms6989 (2015).
Deng, Q. et al. Down‐regulation of tissue N: P ratios in terrestrial plants by elevated CO2. Ecology 96, 3354–3362 (2015).
Lam, S. K., Chen, D., Norton, R., Armstrong, R. & Mosier, A. R. Nitrogen dynamics in grain crop and legume pasture systems under elevated atmospheric carbon dioxide concentration: A meta-analysis. Global Change Biology 18, 2853–2859, https://doi.org/10.1111/j.1365-2486.2012.02758.x (2012).
Jin, J., Tang, C., Armstrong, R., Butterly, C. & Sale, P. Elevated CO2 temporally enhances phosphorus immobilization in the rhizosphere of wheat and chickpea. Plant Soil 368, 315–328, https://doi.org/10.1007/s11104-012-1516-9 (2012).
Luo, Y., Hui, D. & Zhang, D. Elevated CO2 stimulates net accumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology 87, 53–63 (2006).
Cleland, E. E., Chuine, I., Menzel, A., Mooney, H. A. & Schwartz, M. D. Shifting plant phenology in response to global change. Trends in Ecology &. Evolution 22, 357–365 (2007).
Bhattacharyya, P. et al. Effect of elevated carbon dioxide and temperature on phosphorus uptake in tropical flooded rice (Oryza sativa L.). European Journal of Agronomy 53, 28–37, https://doi.org/10.1016/j.eja.2013.10.008 (2014).
Acknowledgements
This work was financially supported by National Key R & D Program of China (2017YFD0300202) and Ministry of Science and Technology of the People’s Republic of China (2012BAC19B01). Construction and maintenance of the experimental system was funded by the state Special Fund for Agro-scientific Research in the Public Interest “Climate Change Impacts on Crop Production and Mitigation” under grant number 200903003. The first author also appreciates the China Scholarship Council for providing funds to him to pursue his study in Japan.
Author information
Authors and Affiliations
Contributions
J.W. carried out the experiment, performed analyses and drafted the manuscript; X.L. and G.P. conceived the idea and led the study design; Q.L. and X.Z. contributed ideas to the study and assisted in analyzing the data; S.L contributed the improvement of our manuscript. All authors approved the final version.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, J., Liu, X., Zhang, X. et al. Changes in plant C, N and P ratios under elevated [CO2] and canopy warming in a rice-winter wheat rotation system. Sci Rep 9, 5424 (2019). https://doi.org/10.1038/s41598-019-41944-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-41944-1
This article is cited by
-
Soil and foliar Si fertilization alters elemental stoichiometry and increases yield of sugarcane cultivars
Scientific Reports (2023)
-
Alteration in biochemical constituents and nutrients partitioning of Asparagus racemosus in response to elevated atmospheric CO2 concentration
Environmental Science and Pollution Research (2022)
-
Mitigation of Water Deficit in Two Cultivars of Panicum maximum by the Application of Silicon
Water, Air, & Soil Pollution (2022)
-
Flooding alleviates copper stress on Salix: Evidence from stoichiometric patterns among plant tissues
Plant and Soil (2022)
-
Effects of elevated CO2 on resistant and susceptible rice cultivar and its primary host, brown planthopper (BPH), Nilaparvata lugens (Stål)
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
