
Abstract
Giardia is an intestinal protozoan parasite that has the ability to infect a wide range of hosts, which can result in the clinical condition ‘giardiasis’. Over the years, experimental research has shown the crucial involvement of IL-17A to steer the protective immune response against Giardia. The development of the protective response, as reflected by a significant drop in cyst secretion, typically takes around 3 to 4 weeks. However, early-life infections often have a more chronic character lasting for several weeks or months. Therefore, the aim of the current study was to investigate the dynamics of a Giardia muris infection and the subsequent host immune response in neonatal mice infected 4 days after birth. The outcome of the study showed that a G. muris infection in pre-weaned mice failed to trigger a protective IL-17A response, which could explain the prolonged course of infection in comparison to older mice. Only after weaning, a protective intestinal immune response started to develop, characterized by an upregulation of IL-17A and Mbl2 and the secretion of parasite-specific IgA.
Similar content being viewed by others
Introduction
Giardia is a highly prevalent intestinal protozoan parasite with a wide vertebrate host range, including humans. An infection with Giardia can either pass asymptomatically or can in some cases lead to the clinical condition of giardiasis, which is characterized by gastro-intestinal complaints such as diarrhoea, abdominal pain, nausea and weight loss1,2.
In recent years, it has become increasingly evident that the development of protective immunity against a Giardia infection is regulated by the IL-17A/IL-17RA axis3. Several studies reported the upregulation of IL-17A during infections in both mice and cattle4,5,6. Furthermore, re-stimulation of CD4+ T cells from human patients that were previously infected with Giardia resulted in an upregulation of IL-177. IL-17A subsequently triggers the production of a range of antimicrobial peptides and complement factors and regulates the secretion of parasite-specific IgA’s in the intestine5,8. The binding of these components to the Giardia trophozoites is thought to be crucial to clear the infection from the intestine. Infection studies, performed in mice, gerbils, cats and dogs4,9,10, show that the development of immunity, as reflected by a significant drop in cyst secretion, typically takes around 3 to 4 weeks. In contrast, other studies describe chronic Giardia infections that last for several weeks to even months in calves6, goats11 and sheep12. Importantly, chronic infections are typically observed following early-life infections in the first days or weeks after birth. For instance in cattle, the prevalence is often higher in pre-weaned calves, with a peak prevalence between 1 and 8 weeks of age13,14, than in weaned calves15,16. Likewise, a high Giardia prevalence in young animals is also common in for instance reindeers17, lambs18, cats and dogs19. In humans, Giardia is also more prevalent in children, with a peak prevalence in infants and pre-school children, and is declining with increasing age20.
These observations suggest that there might be an age dependent effect on the immune development process against this parasite. The aim of the current study was to investigate the dynamics of a G. muris infection and the subsequent host immune response in neonatal mice infected 4 days after birth.
Materials and Methods
Infection experiments and tissue sample collection
All animal experiments were conducted in accordance with the E.U. Animal Welfare Directives and VICH Guidelines for Good Clinical Practice. Ethical approval to conduct the studies was obtained from the Ethical Committee of the Faculty of Veterinary Medicine, Ghent University (approval number EC 2017/97).
The setup of the artificial infection study is schematically shown in Fig. 1. C57Bl/6 pups, both males and females, were collected from 6 different litters. The pups from 3 litters were infected 4 days after birth with 10*3 G. muris cysts suspended in 20 μL phosphate-buffered saline (PBS), administered orally with a pipet. The pups from the other litters remained uninfected and received only PBS and thus served as controls. All pups were weaned at 28 days old. At day 7, day 21 and day 60 post-infection, 5 pups were randomly selected from both the infected and control litters. At the time of necropsy, a 2 cm long duodenal tissue sample was taken from each animal. The samples were snap-frozen in liquid nitrogen and subsequently used for the extraction of total RNA. In short, samples were crushed, resuspended in TRIzol (Invitrogen) and RNA was further purified using the RNeasy minikit (Qiagen), according to the manufacturer’s instructions. This protocol was complemented with the RNase-free DNase set (Qiagen) for the removal of contaminating genomic DNA by on-column DNAse digestion. The quality of the isolated RNA was verified with an Experion automated electrophoresis system (Bio-Rad) and the concentration was measured with a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies).

Study design of the artificial infection experiment in neonatal mice. Male and female pups from 3 litters were infected 4 days after birth with G. muris cysts. At days 7, 21 and 60 post-infection, corresponding with days 11, 25 and 64 after birth, respectively, 5 pups were selected at random from both the infected and control litters. The pups were euthanized, tissue samples were collected and trophozoites present in the small intestine counted. Faecal cyst counts were performed every 2 days from day 21 p.i. onwards.
From day 21 p.i. onwards, cyst excretion was monitored every 2 days as previously described by Dreesen et al.4. At the time of necropsy, the number of trophozoites present in the intestine was counted as previously described by Paerewijck et al.8 In short, the small intestine was removed, starting from 2 cm behind the first part of the duodenum. The segment was placed in PBS and incubated on ice for 20 minutes before counting the trophozoites by using a hemacytometer. Results are expressed as the absolute number of trophozoites present in the small intestine.
Real-time quantitative PCR
The relative mRNA expression levels of several genes in the small intestine of uninfected and infected mice were measured at the different time points by a quantitative Real-Time PCR (qRT-PCR) approach. This was done using the SYBR green master mix (Applied Biosystems) in combination with a StepOnePlus real-time PCR system (Applied Biosystems). The sequences of all primers used are summarized in Table 1. QRT-PCR and normalization of the data, which was based on the housekeeping genes Hprt1, Tbp and Rplpo, was essentially performed as previously described4. Gene transcription levels were evaluated based on fold difference between the different groups.
ELISA
IgA antibody levels in small intestinal tissue were measured by enzyme-linked immunosorbent assays (ELISA) essentially as described in Paerewijck et al.8. Total IgA and Giardia-specific IgA levels were measured in intestinal samples from infected and uninfected C57BL/6 mice at day 7, day 21 and day 60 post-infection. Small intestinal tissue was ground into powder in liquid nitrogen. A water-soluble protein extract was prepared by dissolving the powder in 150 mM PBS. After centrifugation at 16 000 rpm for 15 minutes, the supernatant was frozen at −80 °C until further use. Protein content of the supernatant was measured with the Pierce BCA protein assay (Thermo Scientific). For measurement of total IgA, Maxisorp 96-well plates (Nunc) were coated with 2 μg/ml sheep anti-mouse IgA (Sigma-Aldrich), for 16 h at 4 °C. For measurement of Giardia-specific IgA levels, plates were coated with G. duodenalis trophozoites. Hereto, trophozoites that were kept in axenic culture in modified TYI-S-33 medium, were centrifuged for 5 min at 800 g, re-dissolved and washed 3 times in PBS. Next, they were re-dissolved in 8% formaldehyde solution. Two mL of formaldehyde solution was added to trophozoites derived from 10 mL of culture. Hundred μl of this solution was added to each well of the plates, which were pre-incubated with poly-L-lysine (Sigma-Aldrich). Trophozoites were allowed to attach to the plates for 48 hours and the plates were subsequently dried at 37 °C for 24 hours. After washing with PBS-Tween20 (0,05%) solution (PBST), all plates were blocked with 2% bovine serum albumin (BSA) (Sigma Aldrich) in PBST. After blocking, the plates were incubated for 1 hour at room temperature with the intestinal protein extracts (200 μg/mL). For measurement of total and Giardia-specific IgA levels, the plates were washed and incubated for 1 hour at room temperature with goat anti-mouse IgA-HRP (Sigma-Aldrich) in a 1/1000 dilution in PBST. All reactions were visualised with ABTS in ABTS buffer (Roche). Optical density was measured at 405 nm with a Tecan plate reader (Tecan).
Statistical analysis
Statistical analysis was carried out using GraphPad Prism software. For the A one-way ANOVA followed by a Dunn’s multiple comparison test was used to determine differences both within and between the control and infected groups. A P-value ≤ 0,05 was considered significant.
Results
Dynamics of a G. muris infection in pre-weaned mice
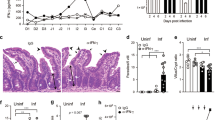
To overcome the technical challenge of collecting individual faecal samples in mice younger than 3 weeks, secretion of cysts in the faeces was only monitored from day 21 post infection (i.e. 25 days of age) onwards. The results of the cyst and trophozoite counts are shown in Fig. 2. While adult mice normally clear an infection with G. muris in approximately 3 weeks’ time21, cyst counts in the neonatally infected mice only started to consistently decrease from day 40 post infection onwards (Fig. 2A). Likewise, trophozoite counts in the neonatally infected mice significantly increased between day 7 and day 21 post infection (i.e. 11 and 25 days of age, respectively), after which they were significantly decreased at day 60 p.i. (Fig. 2B). No significant differences between male and females pups were observed.

Dynamics of a G. muris infection in neonatally infected mice. (A) Cysts present in the faeces of infected mice were monitored every 2 days from day 21 p.i. until day 60 p.i., corresponding with day 25 and day 64 after birth respectively. Mean numbers of cysts per gram faeces obtained from 5 mice at every time point are depicted, with SEM as error bars. The time of weaning, i.e. 28 days after birth, is indicated by a dotted line. (B) The absolute numbers of trophozoites present in the small intestine obtained from 5 infected mice, each represented as one dot with SEM as error bars, are shown at 7, 21, and 60 days p.i. (*p ≤ 0.05, **p ≤ 0.01).
Kinetics of the intestinal immune response following a G. muris infection in pre-weaned mice
The kinetics of the intestinal immune response in the pups was investigated by analysing transcript levels of a range of genes previously shown to be crucial in the protective immune response. These included the cytokine IL-17A, the RAR-related orphan nuclear receptor RORγt which promotes thymocyte differentiation into pro-inflammatory Th17 cells, mannose binding lectin 2 (MBL2) which is a pattern recognition receptor of the innate immune system and the polymeric immunoglobulin receptor (pIgR) which is needed to transport locally produced IgA across mucosal epithelia. The results of the quantitative PCR analyses are summarized in Fig. 3. The results obtained for the negative control mice indicated an age-related expression pattern for some genes. IL-17A transcript was barely detectable at an age of 11 days, but was significantly increased in 25 and 64 days old control pups (Fig. 3A). A similar expression pattern was observed for pIgR (Fig. 3C). Transcript levels of RORγt on the other hand significantly decreased when the pups got older (Fig. 3D). Finally, no significant changes were observed for Mbl2 (Fig. 3B). Comparison between the control and the infected pups showed a significant increase of IL-17A levels at day 60 post infection (i.e. 64 days old), whereas Mbl2 was significantly upregulated at days 21 and 60 post infection (i.e. 25 and 64 days old respectively) (Fig. 3A,B). The transcription patterns of pIgR and RORγt were not impacted by the Giardia infection.

Kinetics of the intestinal immune response following a G. muris infection in neonatal mice. Relative mRNA expression levels of several immune-related genes were measured by qPCR at days 7, 21 and 60 p.i., both in infected and uninfected control mice. Every dot represents data obtained from 1 mouse, with SEM as error bars. (*p ≤ 0.05, **p ≤ 0.01).
In addition to the transcript levels of the above mentioned genes, both the total amount of IgA’s and the levels of parasite specific IgA’s present in the small intestinal tissue were measured by ELISA in both control and infected pups. The results are shown in Fig. 4. In control mice, total intestinal IgA levels significantly increased over time (Fig. 4A). Furthermore, at days 21 and 60 post infection (i.e. 25 and 64 days old), there was a significant increase in total IgA levels in the intestines of infected mice compared to uninfected mice (Fig. 4A). In addition, a significant increase in Giardia-specific IgA’s was observed in the infected mice 60 days post infection (i.e. 64 days old) (Fig. 4B).

Intestinal IgA levels in uninfected control and G. muris infected neonatal mice. (A) Total IgA levels and (B) Giardia-specific IgA levels in the small intestinal tissue were measured by means of ELISA at days 7, 21 and 60 p.i. Optical density is shown in the graphs. Every dot represents data obtained from 1 mouse, with SEM as error bars. (*p ≤ 0.05, **p ≤ 0.01).
Discussion
The outcome of this study shows that a G. muris infection in pre-weaned mice failed to trigger an IL-17A response, which could explain the prolonged course of infection in comparison to older mice. Only after weaning, a protective intestinal immune response started to develop, characterized by an upregulation of IL-17A and Mbl2 and the secretion of parasite-specific IgA. This combination of complement activation and parasite-specific IgA’s has previously been shown to be crucial to combat a Giardia infection8,22,23.
Neonates are generally more susceptible to intestinal infections than adults as their intestinal immune system is still in development following birth24,25. At the time of birth, the intestinal mucosal layer is typically thinner and there are less Paneth cells and immune cells present in the intestinal tissue in comparison with older individuals26. Although Peyer’s patches have already been formed in the murine small intestine at the time of birth, it is only after weaning that they fully mature with the development of germinal centres and activated lymphocytes25. These Peyer’s patches, and in particular the resident macrophages that are present, play an important role in the initial steps of raising an effective immune response following a Giardia infection27,28,29,30,31. In the current study we have shown that the basal IL-17A levels in the intestinal tissue of the neonatal mice were very low. Although the cellular source of IL-17A following a Giardia infection is not yet fully elucidated, at least CD4+ T cells in the intestinal lamina propria seem to play a role5. In the small intestine of neonatal mice, under conditions of homeostasis, CD4+ T cells have an immature phenotype with a reduced synthesis of IL-17A, whereas activated T cells in the lamina propria only seem to be present after weaning32. In the present study, IL-17A transcript levels also significantly increased with age, even in the absence of infection. In contrast, transcript levels of RORγt decreased over time. Although this transcription factor has shown to play a role in Th17 differentiation it is also essential for lymphoid organogenesis, in particular lymph nodes and Peyer’s patches, which could explain its early expression following birth.
In addition to T cells, the presence or absence of antigen presenting cells in the intestinal tract following birth is most likely an important contributing factor. Lantier et al.33 previously showed that the susceptibility of neonatal mice to the protozoan parasite Cryptosporidium parvum was linked to the low number of CD11c + CD103 + dendritic cells present in the gut. Since it was recently shown that the depletion of CD11c + dendritic cells in adult mice impaired the clearance of a G. muris infection34, it would be interesting to also investigate the role of CD11c + CD103 + dendritic cells in the course of a Giardia infection also in neonatal mice. Similarly as in mice, also in calves the number of T cells is significantly lower in newborn calves (1–5 weeks of age) when compared to weaned calves35 and the functional and structural maturation of the Peyer’s patches takes a few months36,37. Above insights, together with the observations made in mice, could explain why the upregulation of IL-17A following a Giardia infection in cattle only becomes detectable after weaning, as previously described4,6.
Following birth, the colonization of the gut by the microbiota is a crucial factor in the full maturation of the intestinal immune system, including the development of lymphoid tissue38,39. For instance, germ-free mice have smaller Peyer’s patches and mesenteric lymph nodes compared to conventional mice40. The bacterial colonization of the gut has also shown to stimulate the expression of the polymeric immunoglobulin receptor (pIgR)41. Also in the present study, an age-dependent upregulation of pIgR was observed in the control mice during a period that coincides with gut microbial colonization. As pIgR is responsible for the transcytosis of soluble dimeric IgA’s inside the intestine, this could also explain the appearance of parasite-specific IgA’s in the intestine following weaning. Interestingly, the administration of broad spectrum antibiotics renders adult mice susceptible to infection with G. duodenalis42, whereas such intervention is not necessary in mice younger than 21 days43. The absence of a fully mature intestinal immune system can explain the susceptibility of neonates, but the mechanisms behind the increased susceptibility of adult mice following antibiotic treatment is still not fully unravelled.
Although neonatal mice seem to be immunologically incapable of clearing a Giardia infection before weaning, ingesting milk from immune mothers was shown to have a protective effect against a G. muris infection44. Similar observations have also been made in both calves and children receiving mother’s milk that contains anti-Giardia antibodies45,46,47. The consumption of such immune milk however does not result in the induction of sustained immunity against this parasitic infection, which implies that the individuals become susceptible again following weaning or when milk consumption decreases44. Nevertheless, reducing Giardia infections in neonates is still important. Recent cohort studies in children have shown that early infections, i.e. in the first 6 months of life, were significantly associated with stunting at an age of 2 years48,49, even when the Giardia infection was not associated with diarrheal symptoms. It was suggested that this could be due to the disruption of epithelial cells by Giardia, however experimental evidence for this hypothesis is still missing. The murine infection model used in this study could be an interesting tool to further investigate the mechanisms behind the long-lasting effects of a neonatal Giardia infection.
References
Geurden, T., Vercruysse, J. & Claerebout, E. Is Giardia a significant pathogen in production animals? Exp. Parasitol. 124, 98–106 (2010).
Heyworth, M. F. Immunological aspects of Giardia infections. Parasite 21, 55 (2014).
Fink, M. Y. & Singer, S. M. The intersection of immune responses, microbiota, and pathogenesis in Giardiasis. Trends Parasitol. 33, 901–913 (2017).
Dreesen, L. et al. Giardia muris infection in mice is associated with a protective interleukin 17A response and induction of peroxisome proliferator-activated receptor alpha. Infect. Immun. 82, 3333–3340 (2014).
Dann, S. M. et al. IL-17A promotes protective IgA responses and expression of other potential effectors against the lumen-dwelling enteric parasite Giardia. Exp. Parasitol. 156, 68–78 (2015).
Grit, G. H. et al. Evaluation of cellular and humoral systemic immune response against Giardia duodenalis infection in cattle. Vet. Parasitol. 202, 145–155 (2014).
Saghaug, C. S. et al. Human memory CD4+ T cell immune responses against Giardia lamblia. Clin. Vaccine Immunol. 23, 11–18 (2016).
Paerewijck, O. et al. Interleukin-17 receptor A (IL-17RA) as a central regulator of the protective immune response against Giardia. Sci. Rep. 7, 1–14 (2017).
Serradell, M. C. et al. Vaccination of domestic animals with a novel oral vaccine prevents Giardia infections, alleviates signs of giardiasis and reduces transmission to humans. npj Vaccines 1, 16018 (2016).
Serradell, M. et al. Cytokines, antibodies and histopathological profiles during giardia infection and variant-specific surface protein-based vaccination. Infect. Immun. 86, 1–12 (2018).
Koudela, B. & Vítovec, J. Experimental giardiasis in goat kids. Vet. Parasitol. 74, 9–18 (1998).
Sweeny, J. P. A. et al. Longitudinal investigation of protozoan parasites in meat lamb farms in southern Western Australia. Prev. Vet. Med. 101, 192–203 (2011).
Geurden, T. et al. A multicentre prevalence study in Europe on Giardia duodenalis in calves, with molecular identification and risk factor analysis. Vet. Parasitol. 190, 383–390 (2012).
Becher, K. A., Robertson, I. D., Fraser, D. M., Palmer, D. G. & Thompson, R. C. A. Molecular epidemiology of Giardia and Cryptosporidium infections in dairy calves originating from three sources in Western Australia. Vet. Parasitol. 123, 1–9 (2004).
Muhid, A., Robertson, I., Ng, J., Yang, R. & Ryan, U. Prevalence of Giardia spp. infection in pre-weaned and weaned calves in relation to management factors. Vet. J. 191, 135–137 (2012).
Xiao, L. & Herd, R. P. Infection pattern of Cryptosporidium and Giardia in calves. Vet. Parasitol. 55, 257–62 (1994).
Niine, T. et al. Giardia and Cryptosporidium infections in neonatal reindeer calves: Relation to the acute phase response. Comp. Immunol. Microbiol. Infect. Dis. 54, 45–50 (2017).
Taylor, M. A., Catchpole, J., Marshall, R. N. & Green, J. Giardiasis in lambs at pasture. Vet Rec 133 (1993).
Barutzki, D. & Schaper, R. Age-dependant prevalence of endoparasites in young dogs and cats up to one year of age. Parasitol. Res. 112, 119–131 (2013).
Polis, M. A., Tuazon, C. U., Alling, D. W. & Talmanis, E. Transmission of Giardia lamblia from a day care center to the community. Am. J. Public Health 76, 1142–1144 (1986).
Roberts-Thomson, I. C., Stevens, D. P., Mahmoud, A. A. & Warren, K. S. Giardiasis in the mouse: an animal model. Gastroenterology 71, 57–61 (1976).
Li, E., Tako, E. A. & Singer, S. M. Complement activation by Giardia duodenalis parasites through the lectin pathway contributes to mast cell responses and parasite control. Infect. Immun. 84, 1092–1099 (2016).
Davids, B. J. et al. Polymeric immunoglobulin receptor in intestinal immune defense against the lumen-dwelling protozoan parasite Giardia. J. Immunol. 177, 6281–6290 (2006).
Stockinger, S., Hornef, M. W. & Chassin, C. Establishment of intestinal homeostasis during the neonatal period. Cell. Mol. Life Sci. 68, 3699–3712 (2011).
Torow, N., Marsland, B. J., Hornef, M. W. & Gollwitzer, E. S. Neonatal mucosal immunology. Mucosal Immunol. 10, 5–17 (2017).
Steege, J. C., Buurman, W. A. & Forget, P. P. The neonatal development of intraepithelial and lamina propria lymphocytes in the murine small intestine. Dev. Immunol. 5, 121–8 (1997).
Heyworth, M. F., Carlson, J. R. & Ermak, T. H. Clearance of Giardia muris infection requires helper/inducer T lymphocytes. J. Exp. Med. 165, 1743–8 (1987).
Carlson, J. R., Heyworth, M. F. & Owen, R. L. Response of Peyer’s patch lymphocyte subsets to Giardia muris infection in BALB/c mice. Cell. Immunol. 97, 44–50 (1986).
Owen, R. L., Allen, C. L. & Stevens, D. P. Phagocytosis of Giardia muris by macrophages in Peyer’s patch epithelium in mice. Infect. Immun. 33, 591–601 (1981).
Hill, D. R. & Pohl, R. Ingestion of Giardia lamblia trophozoites by murine Peyer’s patch macrophages. Infect. Immun. 58, 3202–3207 (1990).
Ermak, T. H., Steger, H. J., Owen, R. L. & Heyworth, M. F. Modulation of lymphocyte subsets in Peyer’s patches of mice treated with monoclonal antibody against helper T-cells. J Histochem Cytochem 36, 417–423 (1988).
Torow, N. et al. Active suppression of intestinal CD4+ TCRαβ + T-lymphocyte maturation during the postnatal period. Nat. Commun. 6 (2015).
Lantier, L. et al. Intestinal CD103+ dendritic cells are key players in the innate immune control of Cryptosporidium parvum Infection in neonatal mice. PLoS Pathog. 9, 1–16 (2013).
Dann, S. M., Le, C. H. Y., Hanson, E. M., Ross, M. C. & Eckmann, L. Giardia infection of the small intestine induces chronic colitis in genetically susceptible hosts. J. Immunol. 201, 548–559 (2018).
Fries, P. N., Popowych, Y. I., Guan, L. L. & Griebel, P. J. Age-related changes in the distribution and frequency of myeloid and T cell populations in the small intestine of calves. Cell. Immunol. 271, 428–437 (2011).
Yasuda, M., Fujino, M., Nasu, T. & Murakami, T. Histological studies on the ontogeny of bovine gut-associated lymphoid tissue: Appearance of T cells and development of IgG + and IgA + cells in lymphoid follicles. Dev. Comp. Immunol. 28, 357–369 (2004).
Tamao, H., Inoshima, Y. & Ishiguro, N. Distribution of immune cells and expression of interleukin receptors in ileal Peyer’s patches of calves. Cell Tissue Res. 346, 245–254 (2011).
Dzidic, M., Boix-Amorós, A., Selma-Royo, M., Mira, A. & Collado, M. Gut Microbiota and Mucosal Immunity in the Neonate. Med. Sci. 6, 56 (2018).
Gensollen, T., Iyer, S. S., Kasper, D. L., Blumberg, R. S. & Medical, H. How colonization by microbiota in early life shapes the immune 1. Gensollen T, Iyer SS, Kasper DL, Blumberg RS, Medical H. How colonization by microbiota in early life shapes the immune system. Science. 2016;352(6285):539-544. https://doi.org/10.1126/science.aad9378. Science 352, 539–544 (2016).
Macpherson, A. J., Köller, Y. & McCoy, K. D. The bilateral responsiveness between intestinal microbes and IgA. Trends Immunol. 36, 460–470 (2015).
Johansen, F., Kaetzel, C. & Genetics, M. Regulation of the polymeric immunoglobulin receptor and IgA transport: New advances in environmental factors that stimulate pIgR expression and its role in mucosal immunity. Mucosal Immunol. 4, 598–602 (2012).
Singer, S. M. & Nash, T. E. The role of normal flora in Giardia lamblia infections in mice. J. Infect. Dis. 181, 1510–1512 (2000).
Hill, D. R., Guerrant, R. L., Pearson, R. D. & Hewlett, E. L. Giardia lamblia infection of suckling mice. J. Infect. Dis. 147, 217–221 (1983).
Andrews, J. S. J. & Hewlett, E. L. Protection against infection with Giardia muris by milk containing antibody to Giardia. J Infect Dis 143, 242–246 (1981).
Mahmud, M. A. et al. Impact of breast-feeding on Giardia lamblia infections in Bilbeis, Egypt. Am. J. Trop. Med. Hyg. 65, 257–260 (2001).
Tellez, A., Winiecka-krusnell, J., Paniagua, M. & Linder, E. Antibodies in mother’s milk protect children against giardiasis. Scand. J. Infect. Dis. 35, 322–325 (2003).
O’Handley, R. M., Ceri, H., Anette, C. & Olson, M. E. Passive immunity and serological immune response in dairy calves associated with natural Giardia duodenalis infections. Vet. Parasitol. 113, 89–98 (2003).
Rogawski, E. T. et al. Use of quantitative molecular diagnostic methods to investigate the effect of enteropathogen infections on linear growth in children in low-resource settings: longitudinal analysis of results from the MAL-ED cohort study. Lancet Glob Heal. 6, (1319–1328 (2018).
Donowitz, J. R. et al. A prospective longitudinal cohort to investigate the effects of early life Giardiasis on growth and all cause diarrhea. Clin. Infect. Dis. 63, 792–797 (2016).
Acknowledgements
This work was supported by the Research Foundation - Flanders.
Author information
Authors and Affiliations
Contributions
Conception or design of the work: O.P., B.M., A.G., K.D.B., P.G. Data collection: O.P., B.M., A.G. Data analysis and interpretation: O.P., B.M., A.G., K.D.B., P.G. Drafting the article: O.P., K.D.B., P.G. Critical revision of the article: O.P., A.G., K.D.B., P.G. Final approval of the version to be published: all authors
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Paerewijck, O., Maertens, B., Gagnaire, A. et al. Delayed development of the protective IL-17A response following a Giardia muris infection in neonatal mice. Sci Rep 9, 8959 (2019). https://doi.org/10.1038/s41598-019-45544-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-45544-x
This article is cited by
-
The NLRP3 inflammasome recognizes alpha-2 and alpha-7.3 giardins and decreases the pathogenicity of Giardia duodenalis in mice
Parasites & Vectors (2023)
-
Regulatory role of the intestinal microbiota in the immune response against Giardia
Scientific Reports (2021)
-
Probiotics in the management of Giardia duodenalis: an update on potential mechanisms and outcomes
Naunyn-Schmiedeberg's Archives of Pharmacology (2021)
-
RORγt+ Treg to Th17 ratios correlate with susceptibility to Giardia infection
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
